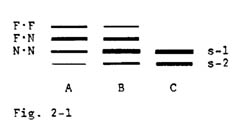
In a line of Papago Flour corn originally obtained from the Maize Cooperation Stock Center, some selfed ears were found to be segregating two additional bands, as in the A and B patterns in Fig. 2-1. In pattern A the faster of these bands stained almost as intensely as the s-1 band, while the intermediate band was strongest. Pattern B was skewed toward the s-1 band. Pattern C contained the usual s-1 band. Upon selfing, patterns A and C were invariant, whereas pattern B segregated all three (A, B and C) patterns. These results are most simply explained by postulating the presence of an additional gene whose product can form a heterodimer with the s-1 band. If the faster (F) and the normally-migrating s-1 band (N) had represented alleles at a single locus, one would have expected segregation of F (alone), pattern A and pattern C in a 1:2:1 ratio in the progeny of a selfed pattern A plant.
Several lines were crossed reciprocally to plants exhibiting pattern A. In nearly all tested cases, only pattern B was observed in the F1 progeny kernels. The fact that the N-N homodimer band was stronger than the F-N heterodimer band in these hybrid plants is consistent with the interpretation that most of the maize lines we tested contain duplicate genes encoding the s-1 band. FN/NN heterozygotes would be expected to exhibit an approximate 9:6:1 relative ratio of the N-N, N-F and F-F bands, respectively, if the two gene products are approximately equal in activity. In all of these kernels there was a positive correlation between the intensity of the s-1 N band and the s-2 band.
In a screening of exotic lines available in our laboratory, a slow-migrating variant (which migrates to the approximate position of the m-1 band) was found to be segregating along with an apparent null for the other s-1 determining locus. The line was characterized by poor germination and general inviability; however, it was possible to recover double homozygous (slow, null) kernels in the F2 generation of an outcross. In order to answer questions about linkage and subunit interactions of the two sMDH genes, plants with this slow, null phenotype were crossed by plants homozygous for the Papago phenotype. The F1 kernels exhibited a five-banded pattern (Fig. 2-2). Upon selfing, six different phenotypes (classified as to number and types of bands) were resolved in the progeny F2 kernels (Fig. 2-2). If the genes were closely linked, one would expect 3 major phenotypes; 25% of the F2 kernels would be "slow, null" and the homozygous FS/FS pattern would be a rare recombinant phenotype. If the genes were unlinked but the slow and fast subunits were alleles of the same locus, one would expect to see the F-F homodimer band segregate out alone 1/16 of the time. If, however, there are two independent genes and the fast and null variants derive from alleles of the same locus, then the ratios in Fig. 2-2 would be expected. Analysis from our initial data from these F2 kernels shows them to be in close agreement to the expected ratios (Fig. 2-2) and thus, the last situation holds. On the other hand, the s-2 band is NOT independent; it varies with the position of the s-1 band (trailing it) and is, therefore, probably due to a modification of one or both sMDH gene products. We see no hybrid bands forming between s-1 and s-2, suggesting that this modification occurs perhaps after dimerization.
Kathleen J. Newton
Return to the MNL 53 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}