Starch gel zymograms of maize malate dehydrogenase (MDH) often reveal complex isozyme patterns. Elucidation of the genetic basis of the observed gel phenotypes has been facilitated by obtaining isozyme variants and analyzing the banding patterns in various combinations. During the last three to four years, such genetic studies have advanced considerably our knowledge of the number and subunit interactions of the maize MDH genes. Based upon independent but concurring evidence from two laboratories (Goodman et al., MNL 52:99, and Newton, these articles), one can propose a model for the inheritance of the cytoplasmic (sMDH) and the mitochondrially-associated (mMDH) forms of MDH.
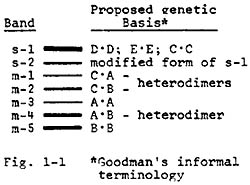
The essential features of this model have been previously suggested by Goodman et al. and are illustrated in Fig. 1-1. Yang's band designations (e.g. in Isozymes, vol. 3) are used in abbreviated form (s-MDH1 is shortened to s-1, m-MDH1 becomes m-1, etc.). They are shown at the left side of the depicted gel banding pattern. The presumed subunit composition of each of the bands is given to the right of the pattern in Goodman's informal terminology.
In most of the commonly-encountered MDH isozyme profiles, the broad, heavily-staining s-1 band includes the homodimeric products of 3 independently segregating genes. Two of these (MdhD and E) encode forms active in the cytoplasm (sMDH's) and the third is a weakly staining mitochondrial isozyme gene (MdhC). The s-2 band appears to be a modified form of the s-1 band. Bands m-3 and m-5 correspond to homodimers of the products of two other independently segregating mitochondrial form genes (MdhA and B), while m-4 is the hybrid band (A-B). Bands m-1 and m-2 also represent heterodimers, between the C and A gene subunits and the C and B ones, respectively. Apart from these five structural loci, a recessive allele of a sixth locus modifies the normal migration of the mitochondrial MDH's. Additional structural loci probably also exist in isolated lines, but most of the patterns we have so far examined can be explained on the basis of the proposed 5 structural gene model.
In the subsequent articles of this series, we report some of the evidence which led us to formulate this type of model as well as results from some localization studies--which support it. In our analyses, slivers of soaked mature scutella were squashed onto filter paper, inserted into 12% starch gels made with a Tris-citrate buffer (pH 7.0) and subjected to electrophoresis for approximately 6 h at 220 volts.
Kathleen J. Newton
Return to the MNL 53 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}