The viviparous mutants of Zea mays are characterized by precocious germination 30-36 days after pollination, indicating a failure of the normal dormancy inducing mechanism. In addition to premature germination, all viviparous mutants cause obvious changes in seed and/or seedling phenotype. These pleiotropic effects allow the identification of homozygous viviparous seed on segregating ears to be made as early as 14 days post pollination.
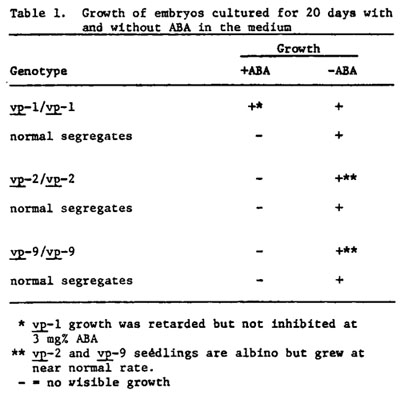
Ears segregating for vp, vp2 and vp9 were harvested at various times from 14 to 30 days post pollination. Embryos were removed from surface sterilized seed and placed on Murashige-Skoog solidified agar medium with and without abscisic acid (ABA). A concentration of 3 mg% ABA was used since this completely inhibited growth of normal embryos. No other growth regulators were added. Explanted embryos were incubated for 20 days at 34C and 100fc illumination in a day:night regime of 12:12. Results are presented in Table 1.
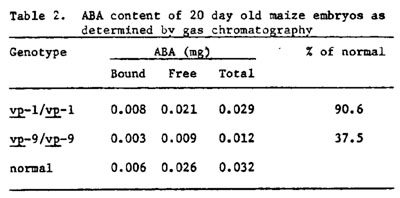
In a second series of experiments, ABA levels of excised 20-day-old embryos were determined by gas chromatography. Preliminary results are shown in Table 2. Homozygous vp seeds were identified by their lack of anthocyanins in full aleurone color genetic background (A A2 C R). Homozygous vp2 and vp9 seeds were identified on segregating ears by their failure to produce endosperm carotenoids in a yellow (Y) genetic background.
Robertson (Genetics 40:745-760, 1955) grouped the viviparous mutants into two classes on the basis of their pleiotropic effects, but our preliminary data suggest that three, perhaps four, classes exist.
Class one mutants have normal carotenoid content in the endosperm and produce normal green seedlings and plants when homozygous. However, they fail to produce anthocyanin in the aleurone in the presence of the genes required for aleurone color. The vp mutant, which belongs to this class, produces normal amounts of ABA, and growth of homozygous vp embryos in culture is not inhibited by ABA at concentrations that prevent growth of normal embryos. We suggest that class one mutants are defective for a specific ABA receptor.
Class two mutants fail to develop carotenoids in endosperm and leaf tissue, and the seedlings are albino when grown under normal illumination. These seedlings are not deficient for chlorophyll, but the chlorophyll is subject to photodestruction in the absence of carotenoids (Anderson and Robertson, Plant Physiol. 35:531-534, 1960). Our preliminary studies with two mutants of this class, vp2 and vp9, show that both are ABA deficient. Embryo ABA content of these mutants is 25-30% of that found in non-viviparous embryos of the same ears at various stages of development tested to date. In addition, the presence of ABA in the culture medium completely inhibits growth of vp2 and vp9 embryo explants. Since ABA and carotenoids have common precursers (Milborrow, Ann. Rev. Plant Physiol. 25:259-307, 1974, and Treharne et al., Phytochemistry 5:581-587, 1966) we suggest that the class two mutants will all be found to be deficient for ABA. The five mutants in this class (vp2, vp5, vp7, vp9 and w3) provide an opportunity to study ABA synthesis using a genetic dissection approach.
Class three mutants have pleiotropic effects on seedlings but do not affect carotenoids or anthocyanins in the seed. Two mutants (vp6 and vp8) fall into this category. While we have no data, as yet, regarding the response of these mutants to ABA, it is consistent with our model to propose that they may affect steps in ABA synthesis beyond the carotenoid branch point.
J. D. Smith, Steve McDaniel and Sam Lively
Return to the MNL 52 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}