In the 1975 Newsletter, occurrence of an aleurone pigment inhibitor which did not give Mendelian ratios was reported. It has since been determined that the plants generating this mutant phenotype had an ancestor from a common genetic stock at some point in their pedigree. In tests involving various sublines of this stock (A C R-g bz), colorless and mottled kernels segregated in the self-pollinated progeny of fully pigmented seeds. When several of the apparently colorless individuals (in a bz bz genotype, small bronze sectors on a colorless background are difficult to detect; therefore, some kernels which appear totally colorless may indeed be faintly mottled with bronze sectors) were selfed, mostly colorless offspring were produced along with a small number of fully pigmented and mottled kernels; there was no apparent ratio. By the second generation of inbreeding, all colorless offspring were obtained, some of which have bred true for two more generations. When a sample of these was testcrossed reciprocally to an r r stock, all progeny were colorless. In crosses of the colorless mutant with recessive stocks of all other aleurone color genes, pigmented offspring were produced. Thus, the mutant is an allele of R.
Among 14 sublines of the A C R-g bz stock tested, 12 generated colorless offspring in varying frequencies; the segregational patterns on the ears indicated that the event responsible for these mutants may be timed in terms of the development of the ear since in a substantial number of cases, the exceptional kernels arose predominantly on the lower half of the cob.
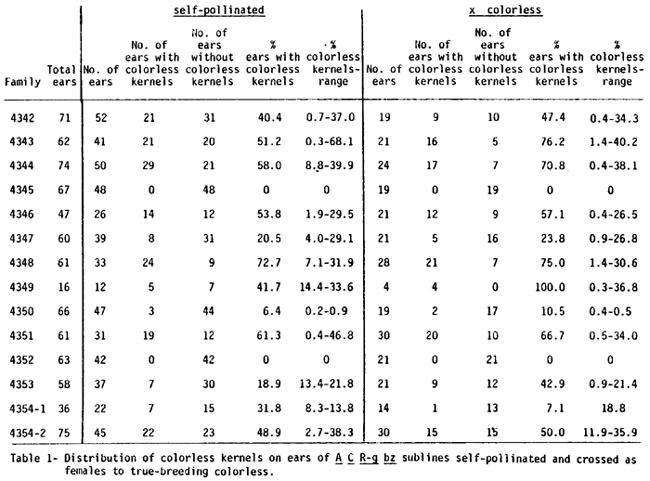
In order to examine the factors involved in generation of these R mutants, sibling plants of each of the 14 sublines were self-pollinated and crossed as females to a true-breeding colorless (mutant) line. Table 1 lists the results. The male parent in these crosses had no apparent effect on either the number of ears segregating for colorless or the frequency of colorless kernels. In family (subline) 4345, neither self-pollinated nor testcrossed ears produced any colorless while at the other extreme in family 4348, about 75% of the ears in each group segregated for the mutant phenotype. Among the remaining families also, the appearance of colorless seemed to be independent of the source of the male gamete.
On those ears which produced mutant kernels, the frequencies ranged from 0.2% to 68.1%. In any given family, there appeared to be no xenia but rather the range of colorless percentages was dependent upon the female parent and was characteristic for a given subline.
In all of the above tests, although the male parents were different, they arose from a common genetic stock. In one case when pollen from an unrelated A C R-r Bz stock was applied to silks of an A C R-g bz subline, the progeny segregated for self-colored, mottled and nearly colorless kernels. Extensive tests, however, have not been conducted to determine the effects of unrelated male parents on the production of colorless kernels in the A C R-g bz sublines.
To determine whether there was an interaction between the newly arisen R mutants and the unrelated R alleles, true-breeding colorless lines were crossed as females with 12 full color stocks and the heterozygous progeny (all self-colored) backcrossed reciprocally to the colorless parent. One of the full color lines used was a W23 stock homozygous for an R-sc allele. On the ears resulting from the backcrosses of colorless females with heterozygous males in all cases except those involving the R-sc allele, there were few self-colored kernels on any ears and the pigmented ones ranged from deeply mottled to nearly colorless. The frequencies of the various color classes varied substantially; on some ears, the deeply mottled kernels comprised nearly 50% of the population while on others, the pigmented class was composed of mostly lightly mottled seeds with a few deeply mottled ones. As yet data on the precise frequencies of each class have not been compiled.
On the ears resulting from the backcross of the R-sc heterozygote as a male to the colorless line, self-colored and colorless kernels segregated in a 1:1 ratio but a few of the offspring on many ears were stippled. From the reciprocal cross of heterozygous females with colorless males, data so far are scant and no firm conclusions can be drawn.
The observations to date suggest that the newly arisen colorless mutant of R is paramutagenic. When it is heterozygous with a number of the R alleles tested, the pigmenting ability of that allele is reduced. The R-sc allele employed is not paramutable in this instance. Whether an inhibitor of R (I-R) is responsible for the few stippled kernels which arose in the backcross progeny of the R-sc heterozygote is not known. Stippled kernels have not been observed in the A C R-g bz stocks although stippling may not be distinguishable from light mottling in a bz bz genotype.
The original event responsible for the mottled and colorless kernels in the A C R-g bz sublines appears to be one which prevents the R allele from completely derepressing in the female. Reciprocal crosses between these stocks and an r tester (and R deficiency) will provide better information regarding this question. Also, whether the R allele which mutates to colorless in tile A C R-g bz stocks is paramutagenic in its pigmenting (derepressed) state or does not become so until after the mutational event has yet to be determined.
J. P. Mottinger
Return to the MNL 52 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}