We have identified and catalogued 855 separate kernel mutants in progenies from material obtained by treatment of pollen with ethyl methanesulfonate according to the method of treatment described below. The development of treatment methods is outlined in recent publications (M. G. Neuffer, 1978, Genetics and Breeding of Maize, D. B. Walden, ed., and M. G. Neuffer and E. H. Coe, 1978, Maydica).
Paraffin oil method for pollen treatment
1. Suspend 0.11% by volume of ethyl methanesulfonate (Eastman #7830) in light domestic paraffin oil (Fisher Lot 722268) by vigorous stirring for an hour or more.
2. Mix EMS suspension with fresh pollen in a plastic vial with a cap, using at least 15 times as much oil as pollen.
3. Shake the pollen-oil mixture periodically to prevent the pollen from clumping in the bottom of the vial.
4. After 50 minutes apply the pollen-oil mixture to fresh silks using a #10 camel hair brush. Apply only enough oil to get a good distribution of pollen. Stirring with the brush is necessary between every application.
Extreme precautions (mixing under a hood, protective clothing, sanitary disposal, etc.) should be taken to protect all persons associated with the work, as chemical mutagens are potentially carcinogenic and are particularly penetrating in oil.
The kernel mutants include a wide range of types from extremely defective mutants to viable mutants such as su and wx. They can be grouped into three broad classes:
1. Those affecting only the endosperm (such as sh, su or wx)
2. Those affecting only the embryo (such as gm)
3. Those affecting both the endosperm and embryo (such as de).
The third class may be divided into (a) viable and (b) lethal types, lethality meaning failure to germinate.
Class 3b and possibly class 2 mutants are of particular interest because they represent a large group of mutants about which very little is known and which hold the promise of including some auxotrophs (Neuffer, 1978). Many of the lethal defective kernel mutants had small, partially-developed embryos indicating that failure was delayed allowing a certain amount of growth. This suggests the possibility that some of these mutants might be rescued by supplying the missing gene product either by genetic manipulation with B-A translocations or by culturing immature embryos on supplemented media.
To identify the proper classes for study, the kernel mutants were screened as indicated in Figure 1.
In the group, 491 mutants incapable of producing viable plants were identified. Of these, 159 were selected for intensive investigation as indicated in the report that follows. A total of 396 kernel mutants including many of the 855 classified above and some from other experiments were tested for location to chromosome arm using J. B. Beckett's series of B-A translocations. A summary of the results is shown in Table 1 (147 mutants located, 37.1%).
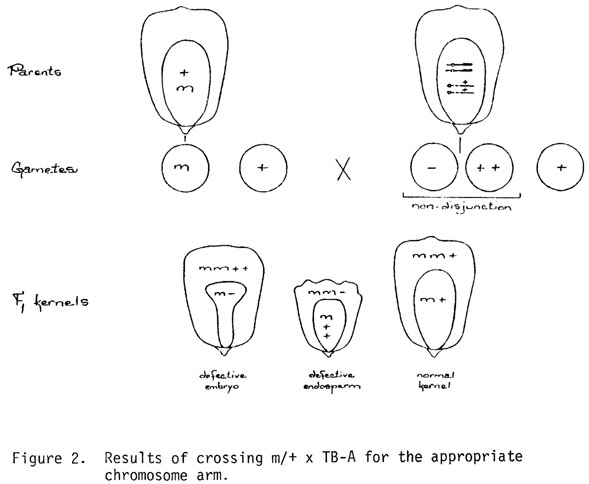
As we proceeded to locate the defective endosperm mutants to chromosome arm we recognized that the TB-A chromosome crosses produced genetic differences in the endosperm and embryo that could be used as a way of rescuing defective kernel mutants with lethal embryos. In the B-A translocation method for locating mutants developed by H. Roman and A. J. Ullstrup (Agron. J. 1951) and perfected by J. B. Beckett (Genetics 1972), each mutant is crossed by a series of B-A translocations in which each of the arms (currently 18 of 20 arms) of the A chromosome set (carrying non-mutant genes) is translocated to the centromere-carrying region of a B chromosome. Non-disjunction of the B centromere at the 2nd microspore division produces sperm nuclei with either 2 copies (duplication) or no copies (deficiency) of the particular chromosome arm involved. If the arm is the one on which the mutant is located, the mutant will be expressed in the cell lineages of the deficient sperm as a hemizygote (m/-).
Assuming a mutant female gamete in the locating cross (+/m x TB-A +/+), the consequences of non-disjunction in the male are such that: (a) the endosperm which receives no copy of the chromosome in question and is therefore mutant (hypoploid, m/m/-) is associated with an embryo that receives 2 copies and is non-mutant (hyperploid, m/+/+); or (b) an alternative in which the endosperm receives 2 copies and is non-mutant (hyperploid, m/m/+/+) and is associated with a mutant embryo receiving no copies of the locating chromosome (hypoploid, m/-), see Figure 2. This sets up a situation where a potentially lethal mutant embryo is associated in the same kernel with a genetically normal endosperm. The question is, will the normal endosperm feed the mutant embryo or vice versa in the reverse situation.
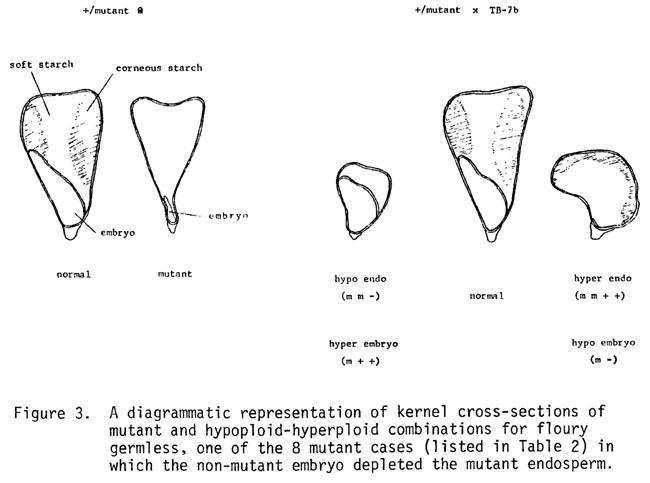
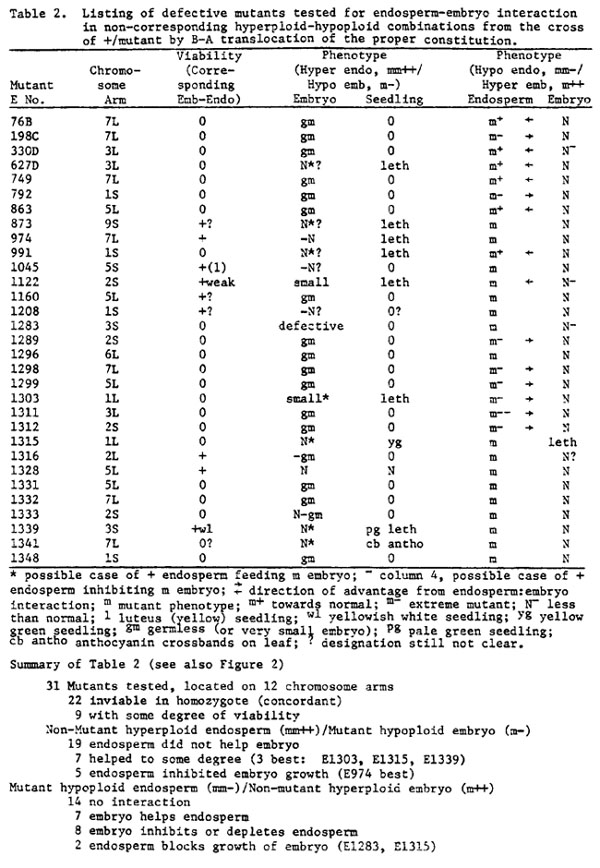
Of the selected 159 mutants, 72 have been located to chromosome arm (45.3%) and of these, 31 have been tested for their response in both embryo-endosperm combinations described above. The results are seen in Table 2.
M. G. Neuffer, W. F. Sheridan and E. Bendbow
Return to the MNL 52 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}