It is known from reassociation experiments that much of the DNA of eukaryotes is highly replicated. If genes are in these replications they will fluctuate in number because of unequal crossing over and unequal sister chromatid exchange. The instability of inbred lines for quantitative characters, which was analyzed by Sprague et al., 1960 (Genetics 45:855-866), could be the result of changes in the number of genes affecting these characters. A population will have chromosomes with a range in the number of units in a replication. An equilibrium is established between the formation of divergent numbers and stabilizing selection pressure, which removes replications with high and low numbers of units from the population. If this selection pressure is less effective in open-pollinating species than in self-pollinating species then we can construct an hypothesis to explain heterosis.
Because quantitative characters are controlled by many genes we must construct a model in which selection acts on a group of different genes which are replicated. This may be best explained by using a crude model. Let us assume that there are three genes (A, B, and C) on different chromosomes that control steps on a synthetic pathway.
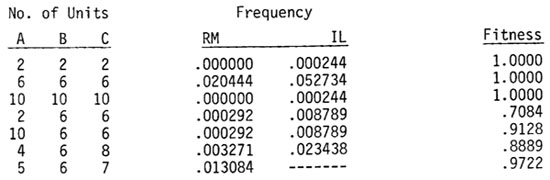
Each of these genes is highly replicated at levels of 1, 2, 3, 4, or 5 (these numbers indicate relative numbers of units). The system is geared for the 3 level (6 in the diploid level). Because of unequal crossing-over or unequal sister chromatid exchange there is divergence from the 3 levels to other levels. The frequency of the different kinds probably approximates a normal curve. It is assumed that their frequencies are: 1(1/16), 2(4/16), 3(6/16), 4(4/16), and 5(1/16)--this is a binomial distribution. There are 53 (125) possible gametes and 93 (729) possible zygotes. Some genotypes found in a randomly mating population and inbred lines derived from it, with their relative fitness, are given below:
The frequency of the various zygotes will be different in a randomly mating population (RM) and in inbred lines (IL) obtained from it. The frequency of 2-2-2 is (1/256)3 in RM and (1/16)3 in IL. Gametic frequencies of the RM give the zygotic frequencies in the IL. The RM population has less genic imbalance because of the possible combinations of low and high levels from two parents. Also some genotypes such as 5-6-7 (which has a high fitness) are not possible in the IL's.
The fitness, the relative amount of progeny produced by an individual, is suggested to depend on the genic balance of the system. This is given an arbitrary, but relative, numerical value by the use of the following formula where the replication levels of different genes in the same system are multiplied and then divided by their mean value raised to the n power, thus:
It will be seen that the fitness of the first three cases in the table is 1. If the system is independent the efficiency depends on genic balance. A 2-2-2 system would "work overtime" and a 10-10-10 system would "stop work early." Other systems are genically unbalanced and inefficient. It is easy to understand why a 2-6-6 system would be in trouble because of a bottleneck effect. A 10-6-6 system, however, should perhaps have no trouble--there will be only an excess of the A product. However, this may disturb the functioning of the B and C genes. If we consider the poor adaptation of trisomes we see that some sort of mechanism like this is acting. Maize trisomes, for example, have reductions in yield from about 20 to over 80 percent.
If we multiply the frequencies of the genotypes by their fitness we can obtain values for the average fitness of the population. The randomly mating population has a fitness of .9416 and the average fitness of the inbred lines obtained from it is only .8787 (the computations required even for this simple model are lengthy and cannot be given here--xeroxed copies will be sent on request). The difference between the two values is significant but not great. If we consider that the total fitness of a plant is the product of the fitness of several systems then this difference is compounded--thus if there are 10 systems, (.9416)10 = .5478 vs. (.8787)10 = .2744.
Inbreeding tends to increase the genic imbalance. The balance may be restored when two inbred lines are crossed (2-2-10 x 10-10-2 = 6-6-6).
It is known that related species may have greatly different amounts of DNA and this has been suggested to mean that they are geared to different levels of replication. Corn Belt corn was produced by crossing Northern Flints and Southern Dents. It is possible that they had different optimum levels. The result of crossing them is suggested by the following:
The variance of the hybrid is increased and there is a greater potential for genic imbalance in the inbred lines extracted from the hybrid than from either parental race.
The testing of this hypothesis is difficult and the mathematical model needs to be refined. This model of heterosis does not exclude the co-existence of other models.
G. G. Doyle
Return to the MNL 52 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
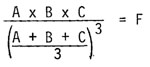{kind=link}
{kind=link}