Nuclear male-sterile genes in hybrid seed corn production may provide useful alternatives to hand detasseling. Additional information on the Duplication-Deficiency (Dp-Df) 5-6b/polymitotic nuclear male-sterility system reported in the 1975 MGCNL (pp. 118-119) supports its utility. Important aspects of the system are stated below followed by the respective documentation.
a) Phenotype of heterozygous Dp-Df 5-6b plants is easily recognized: Plants heterozygous for Dp-Df 5-6b are chromosomally duplicated for 90% of the short arm of chromosome 5. Distinguishing features of the Dp-Df 5-6b heterozygote include shorter and wider leaves; top leaf short, usually wide, stiff, and not bending; tassel branches shorter, upright, and not bending; main tassel spike enlarged in diameter; and generally shorter plant stature. Dp-Df 5-6b heterozygotes shed abundant pollen. These phenotypic features are reminiscent of trisomic chromosome 5.
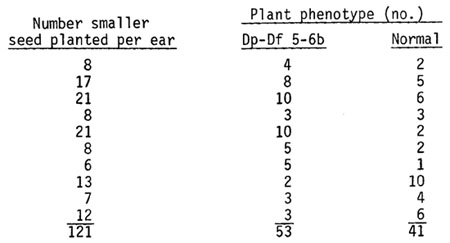
b) Smaller seed selected from the initial cross of heterozygous 5-6b translocation x heterozygous male-sterile (+/po) produces 56% Dp-Df 5-6b plants: Seed about three-fourths the size of the larger kernels was selected from the cross of the 5-6b translocation heterozygote (T5-6b/A632) as the female parent with the heterozygous male-sterile (+/po) stock. The results presented below indicate that 56% of the plants from the smaller seed were Dp-Df 5-6b heterozygotes. The frequency probably would be higher if smaller seed were selected before shelling the ear thus avoiding smaller kernels that occur naturally near the ends of the ear.
Forty plants with the Dp-Df 5-6b phenotype and five with normal phenotype were testcrossed as the male parent onto polymitotic male-sterile plants. Since the male parent in the initial cross was heterozygous male-sterile (+/po) one-half of the resultant Dp-Df 5-6b heterozygotes should carry the po allele and give nearly all male-sterile progeny in the testcross. The half receiving the + allele should give all fertile progeny. Normal plants misclassified as Dp-Df 5-6b types should result in a testcross ratio of 1 fertile:1 male-sterile if the po locus was heterozygous. The results given below indicate that identification of Dp-Df 5-6b heterozygotes was accurate on the basis of plant phenotype alone (no pollen classification). Only one case of misclassification was documented.
In conclusion, selection of smaller seed and the identification of Dp-Df 5-6b heterozygotes on the basis of plant phenotype provides a simple method of extracting Dp-Df 5-6b heterozygotes.
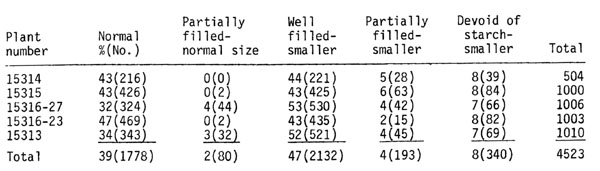
c) Dp-Df 5-6b heterozygotes possess 39% normal, 53% smaller or partially filled with starch and 8% aborted pollen: The various categories and percentages of pollen phenotypes are reported below. Although pollen phenotype can be used to identify Dp-Df heterozygotes, the plant phenotype is sufficiently distinctive.
d) Gametes carrying the Dp-Df 5-6b chromosome are not pollen transmitted: As reported under (e) below, 123 fertile plants have been recovered using the Dp-Df 5-6b/po heterozygote as the male parent. Using a pocket microscope, pollen from each of these plants was examined in the field. In every case the pollen was normal, suggesting that these fertile plants were the result of crossing over in the Dp-Df 5-6b/po male parent and not the result of transmission of a Dp-Df 5-6b chromosome. In addition, the fertile plants always possessed a normal plant phenotype.
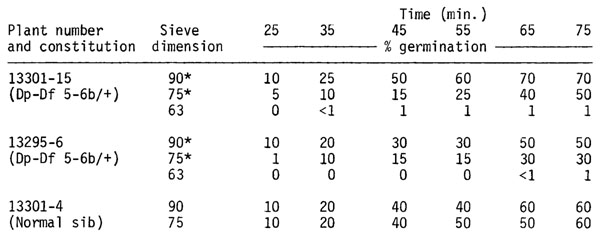
Pollen of Dp-Df 5-6b heterozygotes was sieved through a 125µ/90µ/75µ/63µ/45µ U.S. Standard Sieve series. Pollen collected on the 90µ, 75µ, and 63µ sieves was plated on Cook-Walden solid germination medium. The separation of pollen was incomplete with many smaller grains remaining in the 90µ and 75µ sieves. Direct observation showed that the smaller grains, presumably carrying the Dp-Df 5-6b chromosome, did not germinate. The results reported below indicate that percent germination decreases as average pollen grain size decreases.
The complete lack of male transmission of the Dp-Df 5-6b chromosome as indicated by the normal plant and pollen phenotype of fertile progeny in po po x Dp-Df 5-6b/ po crosses probably can be explained by the effect of the Dp-Df 5-6b chromosome on pollen germination. This nuclear male-sterile system, therefore, is not based on pollen tube competition and should be effective even in environments or with genotypes where there is limited pollen production.
e) At least 98.5% male-sterile plants are produced in male-sterile (po/po) x Dp-Df 5-6b/po crosses: The 1975 MGCNL report indicated that the Dp-Df 5-6b/po had been extracted 8 times yielding 525 male-sterile plants among a total of 526. Additional data were obtained in the summers of 1975, 1976, and 1977. The total number of extractions per year and the number of male-sterile plants among the total tested per extraction are reported below.
The grand total is 8210 male-sterile plants among a total of 8333, or 98.5% male-steriles. Over 58% of the extractions tested yielded all male-sterile plants, based on variable sample sizes.
f) Seed set on the male-sterile (po/po) plants can be genetically improved: A selection program aimed at improving seed set of po/po plants yielded ten lines (5 sister pairs) with good seed set after five alternating self and backcross generations to A632 (ten total generations). These lines traced back to a single selection in the fourth cycle of selection. The material was increased and tested again this season. Several ears on po/po plants were obtained with excellent seed set, suggesting that the improved seed set is heritable. Considerable variability existed among cultures and the number of male-sterile plants tested within a culture was too small to determine the stability of the improved seed set.
g) The Dp-Df 5-6b/po nuclear male-sterility system saves one generation in development: The fact that the male-sterile locus in this system is proximal to the breakpoint (see 1977 MGCNL) allows the extraction of the Dp-Df 5-6b/po heterozygote directly from a cross of heterozygous T5-6b as the female parent with heterozygous po (+/po). If the male-sterile locus is in the duplicated segment (distal to the breakpoint), the female parent must be heterozygous for the translocation and the male-sterile gene, requiring an additional generation. In both situations, only one-half of the resultant progeny will receive the male-sterile allele from the heterozygous po parent and must be identified by a testcross onto the homozygous male-sterile stock.
R. L. Phillips
Return to the MNL 52 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}