We have previously described the isolation and identification of abnormal 10's carrying a duplicate segment in a displaced position (Dp 10-1, -2, -3, -4). Presence of the duplication is inferred from the decrease of recombination values in regions adjacent to the duplication, the abnormal morphology of homozygous Dp10/Dp10 plants and the association of these effects with a gene marker carried on the duplicate segment (the gene component of the R locus, designated P, controlling anthocyanin synthesis in the plant tissues). Linkage studies had shown that duplications Dp10-1, -2, -3 and -4 lie on the long arm of chromosome 10 and in one case (Dp10-1) the cytological observations indicate that the duplicate segment is terminal. The data to be presented refer to additional linkage tests that were performed in order to get more precise information on the physical position of these duplications.
The genotype of Dp10/N10 heterozygotes used to locate Dp 10-1 in relation to R and o7 was R-st +/r-g o7, with the R-st strand carrying the duplication, while in the analysis of Dp10-3 and -4 R-sk +/r-g o7 heterozygotes were used.
The following distribution of seed phenotypes was observed in the progeny of these testcrosses:
aStippled or smoky
bThese data refer to the testcrossed progeny of plants, originally in a Dp 10-3/N1O family, that lost the duplicate segment
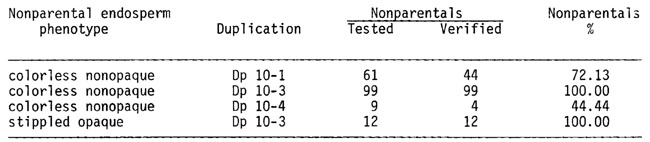
Contrary to expectations, the recovery of the two classes of nonparentals is not equal, the frequency of colorless nonopaque kernels far exceeding that of the stippled opaque ones.
This difference is not likely to be accounted for by the difficulty of detecting the opaque phenotype among the stippled or smoky kernels as proved by the fact that seeds of the latter phenotype are recovered in high frequency in the progeny of heterozygous parents that lost the duplication (see third row in the above table).
Furthermore, progeny tests of both types of nonparentals, even though still incomplete, confirm the validity of the screening method as the data below show:
In the presence of Dp 10-4 phenotypic classification proved to be unreliable. The basis for such a difference in recovery of the two classes of nonparentals remains at the moment unexplained.
Presence of the P marker on the duplicate segment allows the detection of whether the yield of nonparental strands from R-st Dp10 + / r-g N10 o7 parents is associated with loss of the Dp segment.
Data referring to this test, as obtained after progeny tests of presumed recombinants, have been obtained for one kind of nonparentals (colorless nonopaque aleurone) and for Dp 10-1 and -3 only.
These data together with the cytological evidence previously mentioned indicate that Dp 10-1 is most likely distal to o7. The results for Dp 10-3 are best interpreted by assuming the duplication to proximal to o7; if it were distal about 20% (11/56) of the r-g o+ recombinant strands should double crossovers, an unlikely event in view of the positive interference known to occur in the long arm of chromosome 10.
G. Gavazzi and M. Sandri
Return to the MNL 52 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}