Two luteus alleles, oy-1040 and oy-1039 (formerly l*-1040 and l*-1039), derived from D. S. Robertson's mutator line, are lethal and severely defective in the conversion of protoporphyrin IX (proto) to Mg-protoporphyrin (Mg-proto) (Mascia & Robertson, MGCNL 51:38-42, 1977). This is in contrast with the observation that oy is a viable lesion. In an attempt to determine the relationships among these alleles the porphyrins accumulated by these mutants and various combinations have been analyzed.
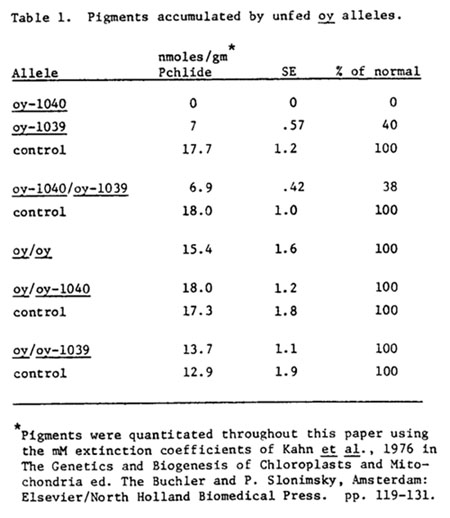
Dark grown plants are normally blocked in the conversion of protochlorophyllide (pchlide) to chlorophyllide (chlide). The pathway is further inhibited by a feedback mechanism preventing d-aminolevulinic acid (ALA) synthesis. Under these conditions normal plants accumulate a small amount of pchlide. Table 1 indicates the pchlide accumulated by oy alleles. Two observations can be made concerning these data. Firstly, oy-1040 is the most stringently blocked allele accumulating no detectable porphyrin pigments, while oy-1039 accumulates approximately 40% of the normal level of pchlide.
Heterozygous oy-1040/oy-1039 seedlings accumulate the same concentration of pchlide as the leaky plants. Since these alleles were derived from closely related sources of the mutator line, the possibility that they arose from a single mutation event with a modifier gene in the leaky stock had to be examined. This can be tested as illustrated in Table 2. If the leakiness of the oy-1039 stock were due to an independent modifier gene, selfs of all of the above combinations would give rise to a 3:1 segregation of leaky to nonleaky seedlings among mutants. Results obtained were consistent with the conclusion that the oy-1039 allele itself is leaky. Secondly, oy and combinations of oy with oy-1040 and oy-1039 were at least equivalent to normal with respect to pchlide synthesis.
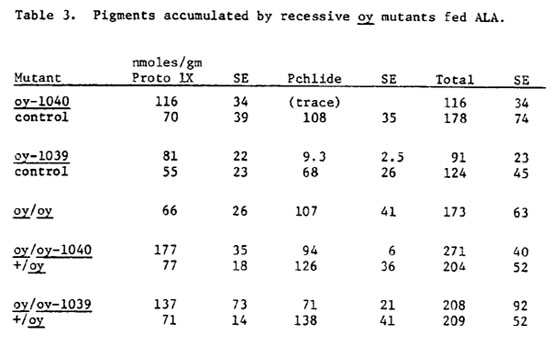
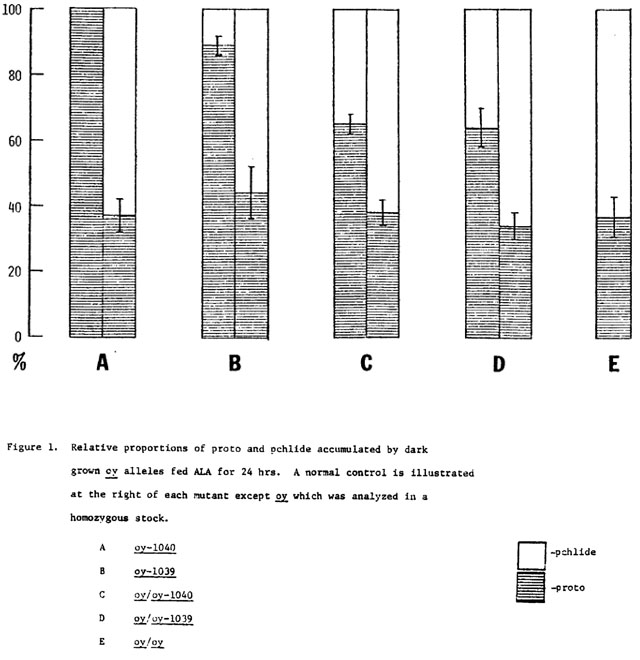
When dark grown plants are fed ALA the regulatory step in chlorophyll biosynthesis is bypassed and significant increases in precursors are accumulated. The actual amount of pigments accumulated by these plants is extremely variable due mainly to variation in uptake of ALA (Table 3). However the proportions of the pigments accumulated are fairly consistent (Figure 1). In these experiments, normal plants accumulated a mixture of approximately 40% proto and 60% pchlide (Figure 1, control values A-D). The most stringently blocked allele, oy-1040 (Figure 1A) accumulates almost exclusively protoporphyrin IX, however, a trace of pchlide may be present in these seedlings. Although slightly leaky, oy-1039 (Figure 1B) accumulates predominantly proto (90%) and only 10% pchlide. It is interesting to note that this mutant does not accumulate significantly more pchlide when fed ALA than when harvested unfed. Homozygous oy plants (Figure 1E) are similar to normal with respect to their precursor accumulation. Heterozygotes between oy and the lethal mutants (Fig. 1C & D) are intermediate in phenotype, accumulating approximately 65% proto and 35% pchlide. The oy allele thus appears to be semi-dominant to the lethal alleles.
It is concluded from these data that the primary function of the oy+ allele is in the conversion of proto to Mg-proto. The two lethal loci reported here, oy-1040 and oy-1039, are severely defective in conversion of proto to Mg proto while oy is sufficiently leaky not to appear defective in this conversion at all. The defect is, however, observed in the plants heterozygous for oy and either of the lethal alleles where one dose of the oy allele is not sufficient to allow normal synthesis of pchlide in dark grown plants fed ALA. Such seedlings, although appearing somewhat green in a seedling bench, are lethal. Several dominant visible, recessive seedling lethal oy alleles, Oy-yg-700 and Oy-yg-1459 have recently been obtained from Dr. Neuffer. Unfortunately, Oy-yg-1459 is in a heterozygous stock and homozygous Oy-yg-700 seeds have failed to germinate. Failure to germinate is also a characteristic of oy-1040 particularly in old seed. It is not unreasonable to speculate that these dominant alleles are also severely blocked in the conversion of proto to Mg-proto. Confirmation of this awaits further testing.
Peter Mascia
Return to the MNL 52 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}