Telocentric B9's carry out nondisjunction at very low rates under most conditions (1-2%) and are considered blocked at some important step in nondisjunction (Chromosoma 42:127). However, in the presence of extra B chromosomes the rate of nondisjunction may rise appreciably. The "leakiness" of telocentric B9's in nondisjunction is examined here.
Data reported earlier for telocentric 1852 (Proc. 1st Int. Maize Genetics and Breeding Conf. - 1975, in press) showed a rise in the rate of nondisjunction from 0.8% to 8% with the addition of 3-8 extra B chromosomes. The rate of B9 instability also increased in the presence of added B chromosomes (Bz/bz sectoring), suggesting that the B9 chromosome was destabilized at the second pollen mitosis by extra B's. The effect of extra B's was either nondisjunction or damage to the B9 centromere, but not restoration of typical B chromosome behavior. It was concluded that telocentric 1852 lacks a cis genetic function essential to nondisjunction.
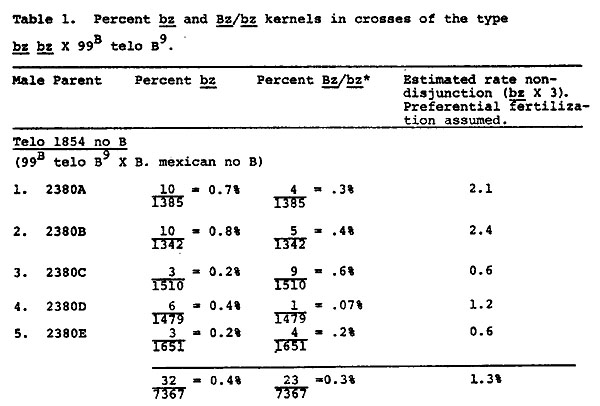
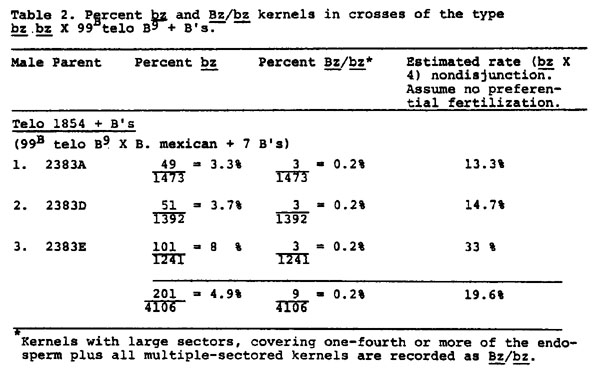
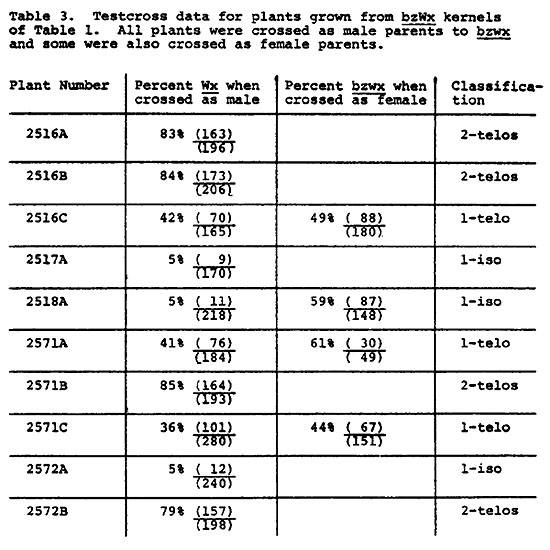
Findings reported here for telocentric 1854 show an increase in the rate of nondisjunction in the presence of extra B's from 1.3% to 19.6% (Tables 1 & 2). The frequency of Bz/bz sectors remains low in both the presence and absence of extra B's. It is difficult to find evidence in the data of Tables 1 and 2 for destabilization of the B9, and one might conclude that extra B's restore to some extent the "normal" nondisjunctional properties of a B chromosome. However, nondisjunction was measured by the expression of the recessive bz phenotype in the endosperm, with the assumption that two telocentric B9's were present in the embryo. To test the assumption, a number of bz kernels from crosses of Tables 1 and 2 were grown and the plants tested for the expected number of two B9 chromosomes. Each plant was crossed as male to a bz wx tester and some plants were also crossed as female. In 9 bz, 9B Wx, B9 Bz, B9 Bz plants, pairing and disjunction of the two B9's produces primarily 9B Wx, B9 Bz and 9 bz wx, B9 Bz meiotic products (D. S. Robertson, Genetics 55:433). Due to the duplicate nature of 9 B9 pollen, the 9 B B9 (Wx) pollen succeeds in fertilization much more frequently than 9 B9 (wx) pollen. The percent of Wx progeny is, therefore, expected to be significantly greater than 50% when two B9's are present. The data of Table 3 indicate that four plants (2516A, 2516B, 2571B, 2572B) from bronze kernels of Table 1 contain two B9's. However, the remaining six plants tested gave rates of Wx transmission less than 50%. Since all six plants show some viability of Wx pollen, they were not missing the B9 chromosome: a 9 wx, 9B Wx plant gives all recessive progeny. It seems apparent that each plant contained one B9. The conclusion was confirmed by reciprocal crosses carried out for some of the plants. For the chromosome type 9 bz wx, 9B Wx, B9 Bz, the frequency of bz wx progeny in a testcross is expected to be quite high in comparison to the low frequency expected with two B9's. A control group, known to carry the telocentric as 9 bz wx, 9B Wx, telo-B9 Bz, gave a bz wx frequency of 56% (1324/2349). Rates of bz wx on the female crosses of 2516C, 2518A, 2571A and 2571C were similarly high. (No data are available for 2517A or 2572A, but their identification is unequivocal, as reported later).
The results indicate that 0-1 disjunction frequently occurs among apparent cases of nondisjunction by telocentrics. However, three of the six plants considered to contain one B9 gave anomalous results in the male cross. Control plants, known to carry 9 wx, 9B Wx telo-B9, gave 47% (1922/4071) Wx progeny when crossed as male parent to a wx tester. Data for 2516C, 2571A and 2571C in Table 3 agree with the control. However, plants 2517A, 2518A and 2572A gave markedly lower frequencies of Wx. Subsequent cytological examination showed that the latter plants contained a B9 isochromosome rather than a telocentric (see accompanying article). Three types of disjunction were, therefore, found: 0-1, 0-2, 0-iso.
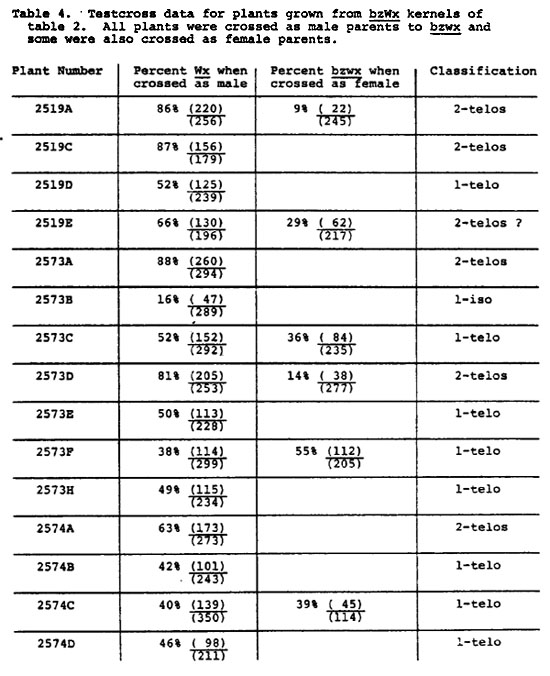
The bronze kernels of Table 2 were also tested for the presence of B9's. Results from the male crosses (Table 4) indicate that 6 plants contained two telo-B9's, 8 plants had one telo-B9 and one plant had a B9 isochromosome. (An additional three plants had two B9's, but these B9's showed very high rates of nondisjunction in testcrosses and probably were standard B9's derived by crossing over between telocentric B9's and standard B chromosomes). There are several difficulties with the data of Table 4. First, the isochromosome classification has not been confirmed cytologically. Second, male transmission of Wx for 2519E and 2574A seems rather low, although well above 50%. Third, the data from female crosses for bz wx frequency do not agree well with the expected in several cases. Among plants believed to have two B9's, 2519A and 2573D have low frequencies of bz wx, as expected. However, 2519E has a very high bz wx frequency, near that expected for one B9. Also, among plants classified as having one B9 from the male crosses, the bz wx frequency on the female cross seems low for 2573C and 2574C. No explanation for the discrepancies can be proposed, but the effect of extra B's on pairing, crossing over and disjunction of B9's may play a role.
Despite the problems outlined, it appears the 0-1 disjunction occurs for telocentric 1854 in the presence of B chromosomes as well as in their absence. Addition of B chromosomes does not eliminate this anomalous behavior at the second pollen mitosis. The finding suggests that B chromosomes contribute to the "stickiness" of the B9 chromosome and make disjunction more difficult. The result may be a rise in nondisjunction, but the extra B's do not restore the missing element(s) of the telocentric which assures migration of the chromatids to one pole without difficulty.
Wayne R. Carlson
Return to the MNL 52 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}