Rhoades and Dempsey have published extensive data demonstrating that the B chromosomes of maize induce chromatin elimination from knobbed chromosomes of the regular complement (A chromosomes). The hi-loss phenomenon takes place at one specific mitosis--the second microspore division. This is the mitosis in which nondisjunction of B chromosomes occurs. Since both phenomena occur in the same cell division, they postulated that the B chromosomes induced a delayed replication of the heterochromatic knobs of the A chromosomes and of the proximal heterochromatic knob immediately adjacent to the centromere of the B. Late replication of the B's proximal knob caused B non-disjunction while delayed replication of the distally situated knobs of the A chromosomes led to the formation and subsequent breakage of the dicentric bridges produced at anaphase.
Their genetic data involving the position and frequencies of chromatin loss for five different knobbed chromosomes are wholly consistent with this hypothesis. However, confirming evidence is needed before giving it unequivocal acceptance. The validity of the Rhoades-Dempsey hypothesis was tested by determining cytologically the kinds of structural changes found in the progeny of hi-loss crosses. To this end, crosses were made with pollen from a hi-loss strain onto silks of yg2 plants. Chromosome 9 of the hi-loss strain used in this experiment had the Yg2 allele, a large knob on 9S and a sufficient number of B's to insure induction of the hi-loss phenomenon. Several chromosomes of the complement were knobbed and these would also undergo breakage and chromatin loss at the second microspore division. The Yg2 locus is near the tip of 9S; all breaks in 9S between Yg2 and the centromere yield a chromosome 9 deficient for the Yg2 allele. Fertilization of the egg pronucleus by this deficient sperm produces a hemizygous yg2 seedling. All of the exceptional yg2 seedlings were grown to maturity and P.M.C.'s taken for pachytene examination. The exceptional yg2 plants which were amenable to cytological analysis all had a deficient 9 as expected. Most of them possessed a paternally-derived chromosome 9 with a simple terminal deficiency but in some plants structural rearrangements were found involving chromosome 9 and another knobbed chromosome. It is through the analysis of these structural rearrangements that critical evidence came bearing on the correctness of their hypothesis.
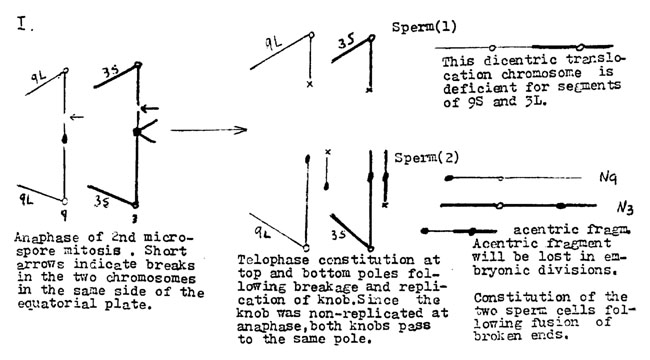
Let us consider the constitution of the sperm cells produced by breakage of the postulated dicentric bridges at the second microspore anaphase following concomitant breaks in the short arm of 9 and in a second knobbed chromosome. For illustrative purposes we have chosen a chromosome 3 with a knob in the long arm. The diagram is self-explanatory.
The Rhoades-Dempsey hypothesis assumes dicentric formation as the result of delayed knob replication. It is further assumed that, following breakage at anaphase, the knobbed acentric fragment passes to the same pole as does the intact knobbed chromatid because the two are conjoined by the still undivided knob. Replication of the knob finally occurs and at telophase or interkinesis the acentric fragment with a freshly-broken end is no longer attached. Fusion of broken ends to give a translocation can occur at this time in telophase nuclei with two broken ends. Presumably it is a matter of chance if breaks in a dicentric bridge occur above or below the equatorial plate.
Sperms 1 and 2 are produced when breaks in the two dicentric bridges are on the same side of the equatorial plate. Sperms 3 and 4 result from one break above and the other below the equatorial plate. Fertilization of a yg2 egg by sperm 1 gives a yg2 plant deficient for segments of chromosomes 3 and 9 and possessing a dicentric chromosome. This chromosome would be subject to strong selection against its survival in the sporophytic tissue because it would undergo the chromosome type of bridge-breakage-fusion and hence tend to be eliminated. It is unlikely that the population of exceptional yg2 plants surviving to maturity would include individuals of this constitution.
Sperm of class 2 would give a normal Yg2 zygote with structurally intact chromosomes 3 and 9.
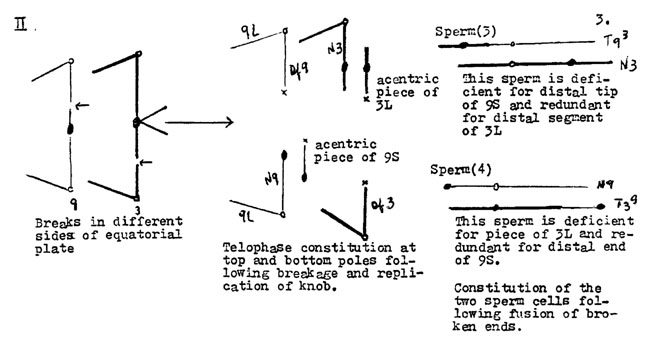
Sperm of class 3 produce yg2 sporophytes with the T93 chromosome (deficient for the Yg2 allele) which is capped by a piece of 3L, a normal chromosome 9 and two normal chromosomes 3. These plants are trisomic for part of 3L and give chains of 4 at diakinesis.
Sperm of class 4 yield Yg2 zygotes with two normal chromosomes 9, one normal 3 and the T39 translocated chromosome.
The zygotes from the functioning of sperms 3 and 4 are complementary. Those from sperm 3 are partial trisomics for chromosome 3 and those from class 4 are partial trisomics for chromosome 9. However, zygotes from class 4 sperm would not be detected in our experiments because they give rise to green plants and loss of the Yg2 allele was the marker used for selecting those plants for cytological studies. They should be found if P.M.C.'s were taken from green as well as yg2 plants.
In summary, when there are concomitant breaks at the second microspore anaphase in two different knobbed chromosomes, the only type of structural rearrangement expected in the exceptional yg2 plants is the class which is partially trisomic. Reciprocal translocations are not expected.
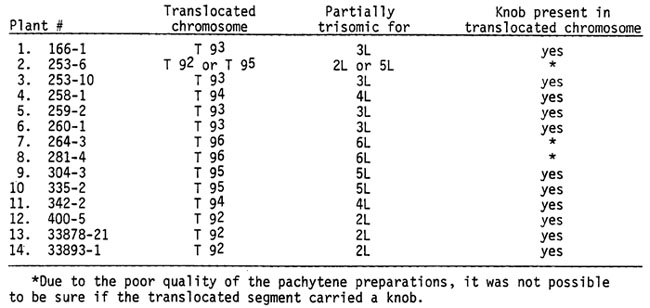
Given above are the theoretical expectations on the Rhoades-Dempsey hypothesis. Do the cytological observations support or negate this hypothesis? To date, I have examined meiosis in 97 exceptional yg2 plants. A simple terminal deficiency in chromosome 9 was found in 83. However, 14 of the yg2 plants possessed a translocation complex derived from breaks in chromosome 9 and in a second chromosome. These are as follows:
Fourteen half-translocations have been generated which involve chromosome 9 and either chromosome 2, 3, 4, 5 or 6, all of which were known to be knobbed in the hi-loss stock. All pieces of a heterologous chromosome translocated to 9S were knobbed.
The reciprocal translocations normally occurring following the induction of breaks in two different chromosomes are non-existent or infrequent among the structural rearrangements found in the progeny of hi-loss plants. Only half-translocations (the recovery of one of the two expected translocated chromosomes) were found although breaks had clearly taken place in two chromosomes. These results are comprehensible on the Rhoades-Dempsey hypothesis, according to which reciprocal translocations should not occur. None were found. Sister sperm 3 and 4 each have one of the two expected translocated chromosomes. However, they were sequestered to opposite poles of the spindle as shown in the diagram and there was no opportunity for the fusion of broken ends needed to produce two reciprocally translocated chromosomes in one nucleus. The finding of 14 half-translocation complexes and of no reciprocal translocations constitute convincing evidence of the correctness of the Rhoades-Dempsey hypothesis.
Luiz Sergio Saraiva*
*Graduate fellow CAPES, Brazil; Permanent address:
Instituto de
Ciencias Biologicas, Universidade Federal de Vicosa,
MG, Brazil
Return to the MNL 52 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}