The Df 33166 chromosome was derived by breakage of a normal chromosome 9 with a large terminal knob on 9S at the second microspore division. Deficient for the Yg2 and Wd loci, its short arm was capped by a small heterochromatic knob of unknown origin. The size of the knob approximated that of the 9S knob present in inbred KYS. Previously reported data demonstrated that the high-loss phenomenon was restricted to knobbed chromosomes in microspores with two or more B chromosomes and that there was a positive correlation between knob size and rate of loss. Since the knob on 33166 was small, it was anticipated that its rate of chromatin loss would be much less than that of KL9. However, data reported by Rhoades and Dempsey in the 1977 Maize News Letter show that: (1) the 33166 chromosome underwent a much higher rate of loss than did the KL9 chromosome in microspores of heterozygous plants, (2) that breaks in the postulated dicentric bridge arising by delayed replication of the small knob of 33166 were predominantly distal to the C locus while the great majority of breaks in the dicentric coming from the KL9 chromosome were proximal to Wx, and (3) that the 33166 chromosome underwent chromatin loss in microspores with 1B chromosome while two or more B's were needed to induce loss in the KL9 chromosome. A summary of the data bearing on the frequency of loss of the C locus with varying numbers of B chromosomes is presented below, for crosses on c c female parents. Items 1, 2, and 3 are from previously reported data; items 2 and 3 are sibs, and items 4 and 5 are sibs.
In line 2 of the above tabulation where the microspores had one or two B's the percentage of mosaic kernels amongst the C class was 543/4573 or 11.9%. All or nearly all of the mosaic kernels came from the bridge-breakage-fusion cycle involving the 33166 chromosome at the 2nd microspore mitosis. In the total of 1628 C kernels listed in line 3, approximately 30% or 488 came from a sperm cell produced by a microspore with one B chromosome (observations by Randolph show that only 30%, instead of the expected 50%, of the microspores from a 1B plant have a B chromosome, because of lagging of the univalent B during meiosis). Breaks distal to C occurred in 63 (12.9%) of the estimated 488 microspores with 1B chromosome. This percentage of chromosome breaks in 1B microspores is very close to the 11.9% C-c mosaicism coming from microspores with 1 and 2B's. In the 1977 Maize News Letter, we suggested that a mechanism other than our hypothesis of a delayed knob replication conceivably could be responsible for the inception of the bridge-breakage-fusion cycle. However, recent cytological observations by Luiz Saraiva (see his report in this News Letter) in this laboratory have demonstrated the validity of the delayed knob replication hypothesis. If we accept the correctness of this mechanism for induction of the bridge-breakage-fusion cycle producing mosaic endosperms, it is apparent that 1B is as effective in producing delayed replication of the small terminal piece of heterochromatin on the 33166 chromosome, which leads to dicentric formation followed by bridge breakage, as are 2B's--i.e., the threshold for inhibition of knob replication is reached by 1B chromosome and adding a second B has no effect. This is in contrast to the behavior of a structurally normal KL9 chromosome where chromatin loss does not occur in 1B microspores and two or more B's are needed. However, it is true for both the KL9 and 33166 chromosomes that once the threshold for delayed replication has been attained there is no enhancement in rate of dicentric formation (and loss) by increasing the number of B's.
The data in lines 4 and 5 demonstrate the stability of the 33166 chromosome in 0B microspores. Both the KL9 and the 33166 chromosome are stable during the megaspore mitoses, irrespective of the number of B chromosomes.
The difference in the location of breakpoints in the dicentric bridges coming from the KL9 and 33166 chromosomes may be ascribed to a weak union of the heterochromatic knob with the euchromatic tip of 9S in the origin of the 33166 chromosome. Under anaphase stress, breakage of the dicentric bridge would preferentially occur at this weak junction. If this be true, the great majority of deficient chromosomes recovered in the embryo should be identical in size.
As a consequence of the rupture of the dicentric bridge, formed by the 33166 chromosome, between the C locus and the knob at the 2nd microspore anaphase, one of the two sperm cells has a deficient chromosome 9 with a freshly broken end and the other has a 33166 chromosome. Following fertilization of the polar nuclei by the sperm with the broken 9, a typical bridge-breakage-fusion cycle is initiated in the first division of the primary endosperm nucleus. The mosaic pattern of colored and colorless aleurone involves all of the aleurone layer, indicating that the original break in the dicentric bridge consistently occurred at the second microspore mitosis.
This pattern was invariably found in the hundreds of mosaic kernels previously examined. However, a 33166 sub-line has arisen in which the mosaic pattern of C and c tissue suggests that the dicentric condition, caused by delayed replication of the terminal heterochromatic knob, may originate at either the second microspore mitosis or frequently in the first endosperm division. Bridge breakage at the first endosperm anaphase gives a cell with a 33166 chromosome carrying the C allele and a sister cell with a broken 9. That fraction of the aleurone layer descending from the cell with a broken 9 will be mosaic for C-c because of the ensuing bridge-breakage-fusion cycle when the break occurs between C and the knob. In such instances, approximately one-half of the kernel will have an aleurone layer uniformly self-colored (no mosaicism) while half will be mosaic for colored and colorless aleurone sectors. The size of the self-colored sector varies as expected from kernel to kernel but the observed pattern of mosaicism clearly indicates that breaks may be induced at the first endosperm mitosis.
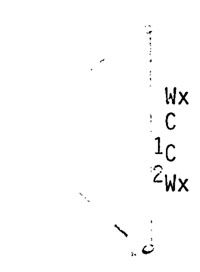
Observations demonstrating dicentric formation at the first endosperm division came from the cross of a c wx tester by pollen from a 33166 C Wx/ N9 c wx plant. The constitution of the dicentric from the 33166 C Wx chromosome produced at either the second microspore or at the first endosperm division is diagrammatically shown below:
Meiotic crossing over between C and Wx gives a 33166 C wx chromosome and a resulting dicentric with the C and wx alleles. Crossovers between C and the knob result in a knob-c-wx chromosome whose involvement in a bridge-breakage-fusion cycle would not be detected genetically. Knobbed c Wx chromosomes capable of causing Wx-wx mosaicism arise from the infrequent double exchanges in the knob-C and C-Wx regions.
From the cross of c wx (Female) X 33166 C Wx/N9 yg2 c wx (Male) the following data were obtained:
The position and time of bridge breakage in the mosaic kernels can be accurately deduced from their mosaic patterns with the exception of the c Wx-wx and c Wx, c Wx-wx classes. Here there is some difficulty in deciding whether the Wx-wx mosaicism includes the whole endosperm or is sectored for stable and mosaic portions. A further difficulty is that the Wx-wx phenotype could represent a break in region 2 between C and Wx in a 33166 chromosome or any break distal to Wx in regions 1 and 2 in a 33166 chromosome of double crossover origin. Even though we know that the great majority of breaks in a 33166 chromosome occur in region 1, we cannot with impunity assign all of the breaks in colorless kernels mosaic for Wx-wx to region 2. Most of them probably are, and are so designated in the tabulation. Considering only the kernels with C-c mosaicism, which come from breaks in region 1, there are 90 from breaks in bridges formed at the 2nd microspore division and 91 from breaks in bridges arising at the first endosperm mitosis. It should be emphasized that the initial breaks at either cell division stem from the delayed replication of the small heterochromatic knob terminating the short arm of the 33166 chromosome. Once the primary break has been induced, the pattern of C-c mosaicism is sensibly similar in endosperms coming from either second microspore or first endosperm breaks--i.e., once the bridge-breakage-fusion cycle has been initiated, the position of subsequent breaks in the developing endosperm is the same in dicentrics of different origin.
An alternative to the above explanation to account for the fractional kernels with half self-colored aleurone and half with the bridge-breakage-fusion cycle is the possibility of a healing in the first endosperm division of the broken end of one of the two chromatids while the other chromatid undergoes the customary bridge-breakage-fusion cycle during development. We are not favorably disposed towards this hypothesis because there is no extant evidence for cessation of the bridge-breakage-fusion cycle during endosperm ontogeny once it has begun.
It is of some interest that Carlson found that nondisjunction of the B9 chromosome was not restricted to the second microspore mitosis and could occur in an early endosperm cell.
M. M. Rhoades and Ellen Dempsey
Return to the MNL 52 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}