An Adh1-S/Adh1-F individual produces three alcohol dehydrogenase (ADH) Set I allozymes: the S-S and F-F homodimers and S-F heterodimers (Figure 1). In starch gels the relative intensities of these three allozyme bands are the reflections of (1) differential expression of the Adh1-S and Adh1-F alleles (D. Schwartz, Genetics 67:411, 1971), and (2) specific activities of the dimers themselves (e.g. Schwartz, TAG 43:117, 1973). A line expressing low levels of an Adh1-S isoallele relative to a standard Adh1-S allele has been described by Y. Efron (Science 170: 751, 1970).
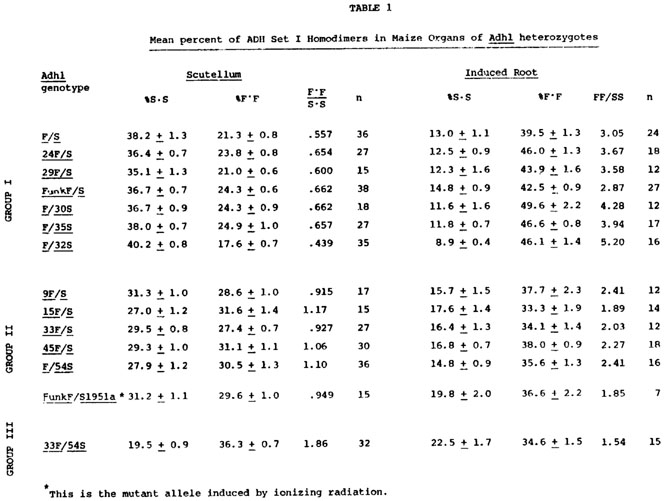
Various heterozygotes were constructed in hopes of detecting ADH regulatory variants on the basis of atypical allozyme ratios. Maize inbreds, exotics and standard Adh1-F/Adh1-S inbreds from lines originated by Schwartz and by Efron were used in these crosses. Two ADH allozyme patterns, one from dry scutellar slices and the other from the induced 5 cm root, were quantified for each heterozygous individual using a Transidyne General TG 2970 integrating densitometer. All the controls are to be published elsewhere. The percent contributions of each homodimer to the total Set I band intensities are reported in Table 1 as mean percent (X) of each homodimer ±95% confidence interval, as calculated from arcsine transformed data. The sources of the Adh1 alleles used in Table 1 are as follows: F and S from Schwartz's inbred F and inbred S lines; 54S from Efron's AdhR-L, Adh1-S line; 9F from Maiz Chapolote; 15F from R2; 24F from Mo. CoB; 29F from Tama Flint; 30S from Parker Flint; 32S from Strawberry Pop; 33F from Super Gold Pop; 35S from Tama Flint Knobless; 45F from R177 and Funk F from Funk G4343 hybrid.
Three distinct scutellar allozyme patterns were found among the Adh1 heterozygotes; the Group II individuals have equal amounts of each homodimer band, whereas the Group I and Group III exhibit unequal amounts of each homodimer band (Table 1). Concurrently, individuals which have the lowest relative amount of a homodimer band in their scutella exhibit the greatest amount of this homodimer in their anaerobically induced primary root (24 hr anaerobic; 5 cm root; see Freeling, MGG 127:215, 1973). The relative amount of the homodimers is expressed as the ratio of the F-F band intensity to S-S band intensity. There is a reciprocal correlation between scutellar F-F:S-S ratio and the induced root F-F:S-S ratio. This reciprocal relationship will be termed "reciprocal effect," and should not be confused with Schwartz's "gene competition" (Genetics 67:411, 1971) or Birchler's "inverse effect" (MGCNL 51:13, 1977). This organ-specific reciprocal behavior is linked tightly to the Adh1 structural gene and is not affected by diverse genetic backgrounds (unpublished results). Since the reciprocal effect segregates with the Adh1 structural gene it may be the property of the structural gene itself or maybe some cis-acting regulatory component of the Adh1 gene. Additionally, since I have measured allozyme balances, some trans effects such as those anticipated on the basis of Schwartz's gene competition hypothesis might occur.
One of the radiation-induced Adh1-S mutants being studied in this laboratory (see following note by Freeling) has an altered reciprocal effect site. Normally, an Adh1-FunkF/Adh1-S heterozygote exhibits a Group I type allozyme pattern. The Adh1-S1951a mutant of this Adh1-S allele, when crossed to Adh1-FunkF, produces heterozygous individuals which exhibit the Group II allozyme pattern in the scutellum and induced root. In short, a mutant can simultaneously lower Adh1 expression in the scutellum while raising expression in the induced root. Ethane methylsulfonate-induced Adh1 mutants have not shown similar behavior. Since Adh1-S1951a is clearly break-point associated (see following note) and since its product is similar to that specified by its Adh1-S progenitor, it seems likely that the reciprocal effect is the property of a site outside of the Adh1 structural gene. Moreover, the variation in ADH Set I allozyme patterns detected among the lines could well be the result of regulatory gene variation. The test of these hypotheses will require careful protein and peptide comparisons, proof of genetic separability, or both.
Perhaps the most important conclusion to be drawn from the data as a whole involves the genetic basis of organ specific differential gene expression. Because a mutant affects Adh1 expression in two cell-types reciprocally, the DNA sequences used to regulate Adh1 in both the scutellum and root must be close-together or identical, and functionally related.
James C. Woodman
Return to the MNL 52 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}