Genetic studies of mitochondrial enzymes open up a new way to approach the possible nature of the so-called "mitochondrial complementation" that attracts particular interest in relation to the problem of heterosis in crop plants (cf. Hanson et al., Crop Sci. 15:62, 1975).
A collection comprising about 50 inbreds and 12 races of maize obtained from Krasnodar Agricultural Research Institute was used in our studies of two enzymes localized exclusively in mitochondria: matrix enzyme glutamate dehydrogenase (GDH) and succinate dehydrogenase (SDH) bound to the inner membrane. The GDH and SDH spectra were investigated primarily in the scutella of three-day-old seedlings, employing the routine procedures of disc-PAGE and tetrazolium staining (in the case of SDH, mitochondria-enriched 10,000xg sediment was used instead of total homogenate).
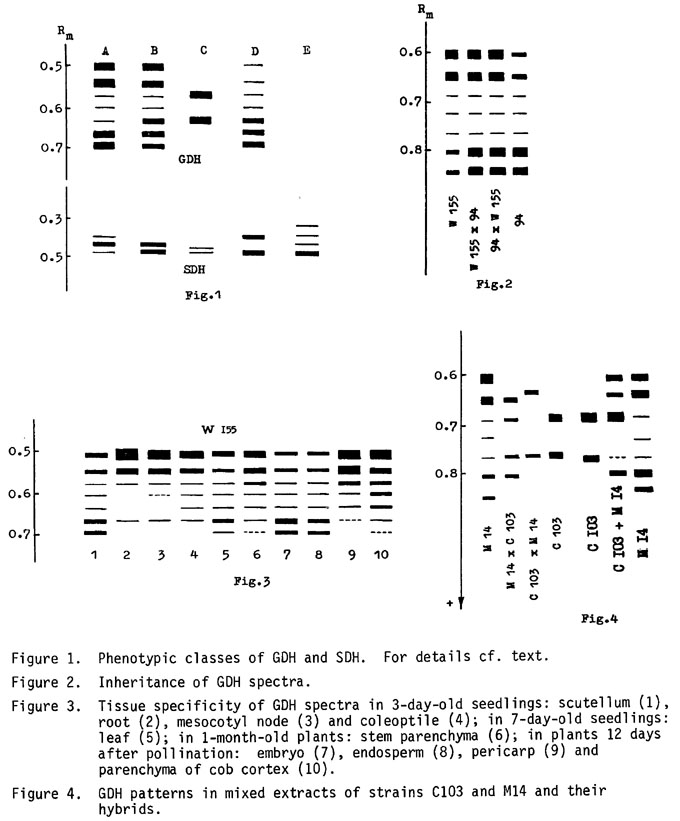
High resolution of GDH spectra (Fig. 1) was achieved using the alcohol dehydrogenase Adh-F band as an inner marker (Rm values were corrected in the case of Adh-S inbreds A seven-band spectrum is characteristic of most of these genotypes, and two classes (A and B) differ only in the staining intensity of the faster bands. An inbred, C103, constitutes class C. Class A occurs with a frequency of about 2/3. Specific differences in staining of the isozymes are genetically determined and are not related to variability dependent on growth conditions. Additive inheritance of isoform staining intensity in reciprocal F1 progenies suggests nuclear control of GDH synthesis (Fig. 2). Log Rm values of these isozymes depend in a linear mode on gel concentration, which means that the seven isozymes of the same molecular weight differ in their charge values. These data are consistent with a two-loci-control model of hexameric GDH (Pahlich, Planta 104:21, 1972; Pryor, Heredity 32:397, 1974). However, the following data do not fit into this model: (1) staining intensity distribution significantly different from binomial predicted by a two-loci model; (2) organ-specific features in GDH spectra (Fig. 3); (3) the spectra of mixed extracts of strain C103 and any class A or B inbred or their hybrids (Fig. 4). These data suggest the presence of some inheritable factor responsible for highly ordered association of GDH subunits and presume the existence of a third locus coding for this factor. Seven- and two-band patterns were maintained after removal of low-molecular-weight substances and partial GDH purification.
Five phenotypic classes of SDH are composed of two to four isozymes controlled by non-allelic genes (Fig. 1); class B occurs at a frequency of about 2/3. Isozymes are of the same molecular weight and differ in their charge values. No tissue- and organ-specific differences have been found in isozyme patterns. Additive spectra of reciprocal hybrids (Fig. 5) suggest nuclear control of SDH. The simultaneous appearance of two SDH classes, A and B, in the Jalisco group 33 race from Mexico and the Morocho race from Argentina is especially interesting as an apparent example of the development of genetic polymorphism in mitochondria.
No new patterns of GDH and SDH were found in the collection of races representing most of the presumed centers of maize domestication, and the frequency distributions of phenotypic classes were similar in collections of races and inbreds.
L. E. Monastyreva, T. B. Sukhorgevskaya and E. E. Khavkin
Return to the MNL 51 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}