The high chlorophyll fluorescence phenotype in maize has been used to
select plants blocked in light reaction photosynthesis. High fluorescent
plants were selected from M2 material ultimately derived from chemically
treated pollen and normal silks as described in MGNL 45:146. Our material
was recovered by screening sandbench material at night using a hand-held
UV lamp (UVL 56, Ultra-Violet Products, Inc.) to excite fluorescence and
a pair of welder's goggles fitted with UV-protective filters to view the
characteristic red chlorophyll fluorescence. Individuals marked at night
were taken from the bench the next day and the hcf phenotype confirmed
by measuring whole-leaf fluorescence induction kinetics in a fluorimeter.
Seed was obtained from families segregating confirmed hcf individuals and
the phenotype examined under growth chamber conditions. The material flowed
through our hands in the following manner:
| No. families | % Total | |
| M2 families screened | 3191 | 100 |
| Number of families segregating hcf in the greenhouse | 155 | 4.0 |
| hcf kinetics confirmed in lab-sandbench material | 105 | 2.7 |
| hcf kinetics confirmed in growth chamber material | 57 | 1.5 |
In preliminary tests, all segregated 3:1 in selfs.
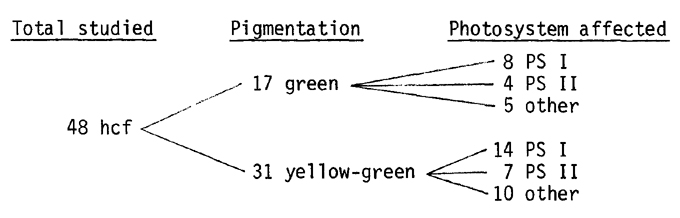
Of these 57 families, 48 have been examined in some detail. On the basis of standard biochemical and biophysical tests the lesions occurring in these mutants can be classified as affecting either photosystem I or photosystem II preferentially. In some cases the site affected by the lesion is not at all clear. The data presently available allow us to draw the following profile:
It is clear that the hcf phenotype is independent of the amount of chlorophyll present in mutant leaves. During the initial selection phase many families segregated YG plants which did not appear to fluoresce highly.
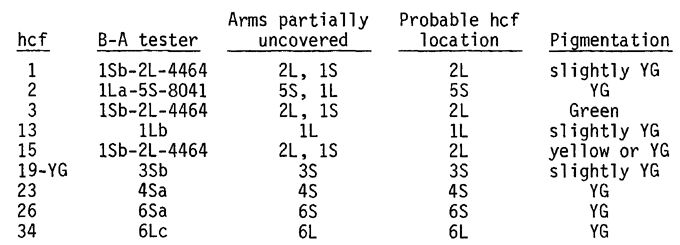
As seed became available an attempt was made to locate the hcf markers to chromosome arm following the procedures outlined in MGNL 45:144; 46:130; and 47:148 using an improved set of B-A translocations. We have been able to locate tentatively nine hcf mutations to chromosome arm as listed below (terminology after Beckett, MGNL 50:89):
In cases where the compound translocations of F. A. Rakha and D. S. Robertson (Genetics 65:223, 1970) or Robertson (MGNL 49:79) were used, the B-A translocations used to develop the compound tester were also used as male in the mapping test and failed to uncover fluorescing individuals. We are reasonably certain, therefore, that the markers lie on the designated arms although further confirmation is needed.
We now know the electron carriers or biosynthetic
capacities affected by mutation in some of our lines (previous article).
Coupling this information with the tentative map locations, it was possible
to select from the larger collection individuals that might be related
genetically. The following allelism tests were performed:
| Same arm: | Missing cyt. b559: |
| 15 x 1 not allelic | 9 x 19 not allelic |
| 15 x 3 not allelic | 9 x 3 allelic |
| 1 x 3 test failed | 19 x 3 test failed |
The other obvious test, that involving hcf-2, 6, and 38 (all missing cyt. f) was carried out but gave ambiguous results in the first test.
The interesting finding is the allelism between hcf-3 and 9. We originally assumed that hcf-3 and hcf-9 might not be allelic since, while both are missing Q and cyt. b559HP, hcf-9 alone is missing both forms (HP and LP) of the cytochrome. The finding that hcf-3 and hcf-9 are allelic raises some interesting possibilities; for instance, we may hypothesize that a single protein necessary for photosystem II function has been altered at two different sites by mutation such that 1) in both cases (hcf-3 and 9) the protein is non-functional as an electron carrier and hence seedlings are hcf; 2) it is unable to bind or allow the proper integration of either form of the cytochrome into the thylakoid membrane in the case of hcf-9; and 3) it can bind or create a favorable membrane environment for the proper integration of b559LP but not b559HP in the case of hcf-3. This is of course only one possible hypothesis, but it is consistent with experimental observations made on these two mutants.
Although the allelism tests for hcf-19 x hcf-3 failed due to poor field conditions it is improbable that at least the yellow-green form of hcf-19 is allelic to hcf-3 since hcf-19YG appears to map to the short arm of chromosome 3 while hcf-3 tentatively maps to the long arm of chromosome 2. Excluding confounding factors (suppressors, background effects) the following conclusions can be drawn at this time: 1) hcf expression is independent of the amount of chlorophyll present (although extremely YG or yellow plants are obviously deficient in the fluorescing species); 2) hcf-3, hcf-9 and hcf-19 are pleiotropic mutants affecting photosystem II function; hcf-3 and hcf-9 are allelic although their biochemical properties differ slightly; both are probably not allelic to hcf-19 YG and possibly not allelic to hcf 19 green; 3) A gene coding for NADPH diaphorase is located on the long arm of chromosome 2; 4) Subject to confirmation an hcf marker, (which is also conveniently yellow-green) is located on the short arm of chromosome 6 beyond the NOR. (Partially supported by NSF Grant PCM 76-08831 and the University of Missouri Research Council). We wish to thank M. G. Neuffer, E. Coe, and J. Beckett for advice, material, and field facilities used during this study.
Kenneth Leto and Donald Miles
Return to the MNL 51 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
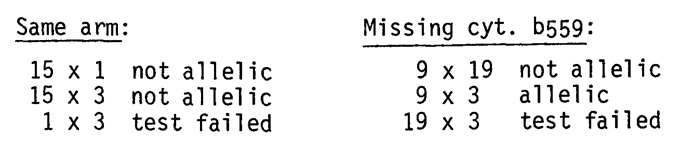{kind=link}