We are currently using high fluorescent mutants in maize to study the mechanism of light reaction photosynthesis. Although photosynthetic mutants have been used to advantage in algae (Levine 1969, Ann. Rev. Plant Physiol. 20:523), very few large scale systematic studies of photosynthesis have been carried out using the mutant approach in higher plants. To date we have obtained about 70 isolates blocked in photosynthesis by selecting high fluorescent seedlings from M2 material as described previously (MGNL 46:127).
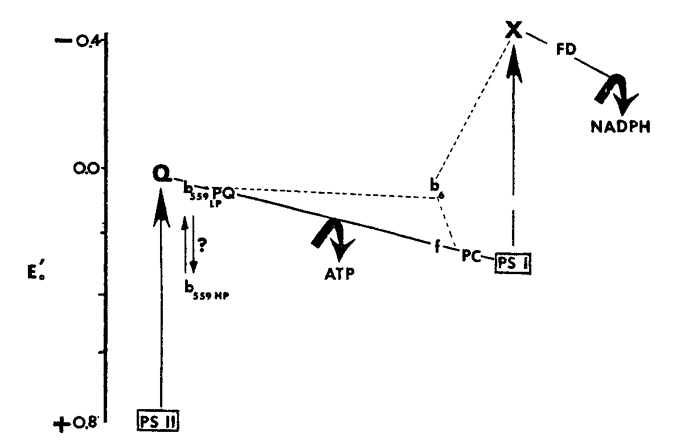
The high level of chlorophyll fluorescence (hcf) seen in mutant seedlings is a result of the inability of these plants to carry out normal rates of electron transport. The light-induced flow of electrons in chloroplasts is mediated by a chain of electron carriers whose arrangement with respect to a chemical potential gradient can be represented schematically by the familiar Z-scheme originally formulated by Hill and Bendall (1960):
Normally, light trapped by photosystem II (PS II) and photosystem I (PS I) reaction centers induces electron flow and the resulting chemical energy is efficiently used in the production of ATP and NADPH. Blockages in electron flow due to missing electron carriers or inefficient production of ATP and/or NADPH result in an inability to dissipate trapped light energy in useful ways and elevated levels of chlorophyll fluorescence result. This phenomenon is independent of the level of pigmentation.
During the past few years we have been able to identify by various means some of the electron carriers and biosynthetic processes affected by mutation in our stocks, as follows:
hcf-1: (C. D. Miles and D. J. Daniel 1974, Plant Physiol. 53:589) reduction in NADPH diaphorase activity; slightly yellow-green (YG) semi-lethal (survives under good field conditions with poor or little seed set).
hcf-2: (Miles and Daniel 1974) loss of cytochrome f and most of the plastoquinone pool (PQ); YG; seedling lethal.
hcf-3: (Miles and Daniel 1974) loss of Q (physiological quencher of chlorophyll fluorescence, presumably a protein), loss of cyt b559 high potential (b559HP); fully green; seedling lethal.
hcf-4: reduced capacity for ATP synthesis, over half PQ missing; slightly YG; semi-lethal.
hcf-6: loss of cyt. f; slightly YG; seedling lethal.
hcf-9: loss of Q and both b559HP and b559LP; fully green; seedling lethal.
hcf-47: missing cyt. b6; YG; seedling lethal.
hcf-48: greatly reduced capacity for ATP synthesis; YG; seedling lethal.
In all cases except hcf-19, segregation is as a single gene nuclear mutation giving 3:1 ratios in selfs and F2 populations (hcf-47 not yet carried to F2). The green:yellow-green heterogeneity present in family hcf-19 has only recently been detected and is not yet genetically investigated. Selfed material segregates approximately 1:1:2 normal but independence has not been established.
It is apparent that several of the mutations are pleiotropic, notably hcf-3, 9 and 19. This observation has its counterpart in the mutational analysis of algal photosynthesis, where the loss of a 47K polypeptide has been correlated with the simultaneous loss of both Q and cyt. b559 in Chlamydomonas reinhardtii (Chua and Bennoun 1975, PNAS 72:2175). From this work and that of other investigators in the field, it appears probable that at least some of the proteins present in the thylakoid membrane are necessary for binding electron carriers.
(Partially supported by NSF Grant PCM 76-08831 and the University of Missouri Research Council). We wish to acknowledge gratefully M. G. Neuffer as the source of the M2 material used in this study.
Kenneth Leto and Donald Miles
Return to the MNL 51 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}