The standard genetic map for maize places ragged (rgd) at the zero position of the chromosome 6 linkage map and polymitotic (po) at the 4 position. The genes are both shown as proximal to the nucleolus organizer region (NOR). The proximal location is suggested by the fact that Kramer and Duclos (MGCNL 36:66) mapped po proximal to rgd from three-point F2 data, and rgd was found by Palmer and Dempsey (MGCNL 42:75) not to be uncovered by TB-6a (breakpoint midway in NOR-heterochromatin) or by T2-6(5419), T4-6(4341), T6-9a or T6-9(4778). All of these interchanges have a break in the NOR.
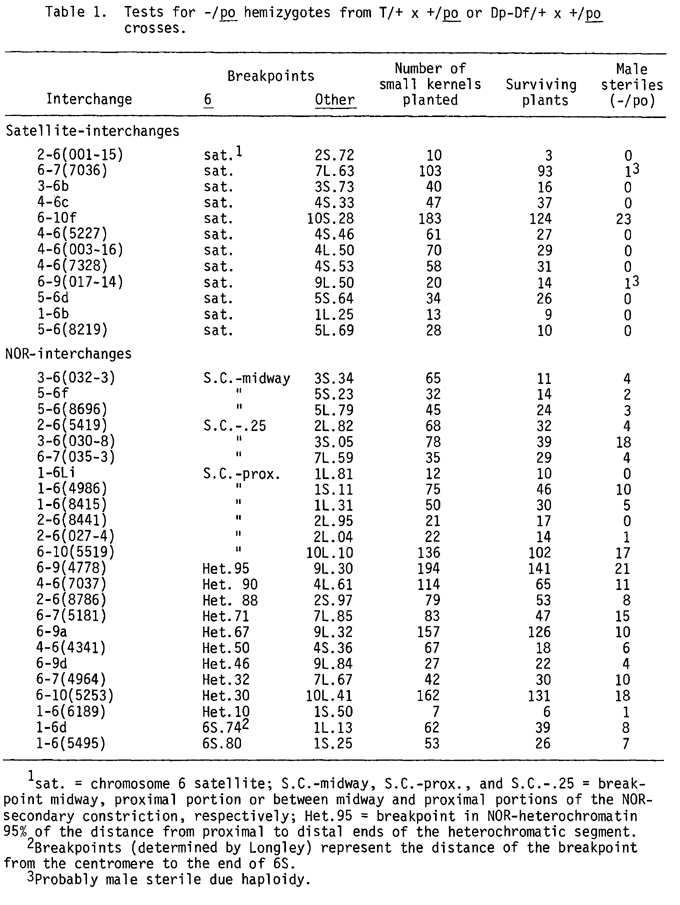
The data reported here place po either in the first chromomere of the satellite of chromosome 6 or in the distal half of the site giving rise to the NOR-secondary constriction. The rgd locus is placed either as proximal to the NOR or in the proximal 10% of the NOR-heterochromatin. The evidence comes from two approaches. First, 10 of the 13 available interchanges with a break in the satellite have been tested for their usefulness in the duplicate-deficient (Dp-Df) scheme of utilizing nuclear male-sterile genes (MGCNL 49:118). The useful interchanges are those which have a chromosome 6 breakpoint distal to po; these interchanges have been shown to be T3-6b, T4-6c, T4-6(5227), T5-6b, T5-6d-and T5-6(8219). Limited data also suggest that T2-6(001-15), Tr-6(003-16) and T6-7(7036) are useful. Plants with Dp-Df complements and heterozygous for po have not been recovered from T6-10f.
The above results are consistent with the second approach of placing po by hemizygous tests. These tests consist of crossing the heterozygous interchange (or in a few cases the heterozygous Dp-Df/+) with a heterozygous +/po male parent and scoring male-sterile (-/po) plants. Table 1 gives the results of planting the smaller seeds from such crosses and noting the existence of male-sterile plants; experience suggests that the selection of smaller seeds enriches for duplicate-deficient heterozygotes.
Note that none of the satellite-interchanges listed in Table 1 uncovers po except T6-10f. This result suggests that all of the satellite-interchanges have a breakpoint distal to the po locus except for T6-10f. The cytological placement of the breaks in the three-chromomere satellites of these interchange stocks is reported by Phillips and Wang (this Newsletter). The genetic and cytological data suggest that po is not distal to the first satellite chromomere. An apparent inconsistency is the cytological location of the T6-10f breakpoint in the last satellite chromomere while po is uncovered in crosses of T6-10f heterozygotes x +/po heterozygotes. Pachytene analysis of T6-10f was repeated and clearly showed the break position as reported. Presumably, the T6-10f stock possesses a complicated aberration not fully understood.
The interchanges with a break in the NOR nearly all generate Dp-Df complements that allow the expression of po (Table 1). The only exceptions seem to be T1-6Li and T2-6(8441), which may simply reflect insufficient sample size. These data are consistent with the placement of po either in the distal half of the site giving rise to the NOR-secondary constriction or in the first satellite chromomere. The results also demonstrate that deficiencies for various portions of the NOR are ovule-transmissible. The deficiency can include the NOR-secondary constriction and as much as 90% of the NOR-heterochromatin [see data for T1-6(6189)].
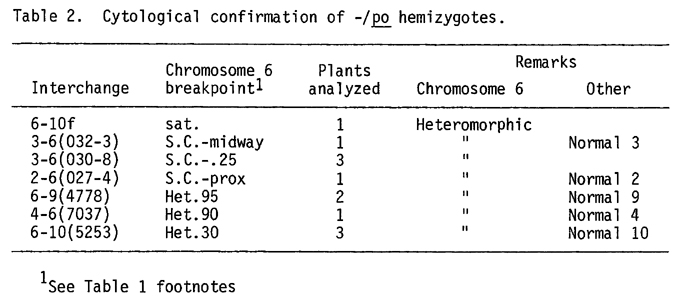
In 1972, Perkins, Newmeyer and Turner (Genetics 71:s46) reported that Dp-Df chromosomes produced from Neurospora interchanges with one break near the chromosome tip could break, resulting in a modified chromosome. Pachytene analyses were performed on many of the putative hemizygous -/po plants in this study to determine (1) if a heteromorphic chromosome 6 bivalent was present as expected in a Dp-Df heterozygote, or (2) if a modified deficient chromosome was present as the result of chromosome breakage. The male steriles chosen for analysis included one from the T6-10f cross, the only satellite interchange that uncovered po, and several NOR-interchanges (Table 2). In many cases, the other chromosome involved in the parental interchange was identified at pachytene; a normal bivalent would be expected and was observed.
In all cases, the expected heteromorphic bivalent associated with the nucleolus was present, indicating that chromosome breakage had not occurred and that the male-sterile plants were hemizygotes for the po locus. They possessed heterozygous deficiencies for parts of the satellite or NOR, depending on the interchange.
With the indication that po is distal to the NOR-heterochromatin, the possibility seemed unlikely that rgd could be distal to po by 4 map units. Therefore, similar hemizygous tests were performed to determine whether interchanges used in this study would uncover rgd. Smaller seeds from crosses of interchange heterozygotes with heterozygous +/rgd plants were germinated in laboratory trays; remnant seed was germinated in a greenhouse sandbench. The lack of ragged plants (Table 3) suggests that rgd is not distal to po but is located proximally either in the short arm of chromosome 6 between the centromere and the NOR or in the proximal 10% of the NOR-heterochromatin. The two markers rgd and po may serve as flanking markers for most if not all of the NOR.
R. L. Phillips, E. B. Patterson and P. J. Buescher
Return to the MNL 51 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}