Response of an olive necrotic mutant to gibberellic acid
Conditional lethal mutants are an important genetic tool for investigating the links between the lethal phenotype and the primary event at the gene level. They might be recovered, in the flowering plants, among those mutants, generally defined as seedling lethals, that die at an early developmental stage when the endosperm nutrients have been exhausted. Their detection rests on the possibility of inducing phenotypic repair by changing environmental factors like temperature, pH, or nutrients.
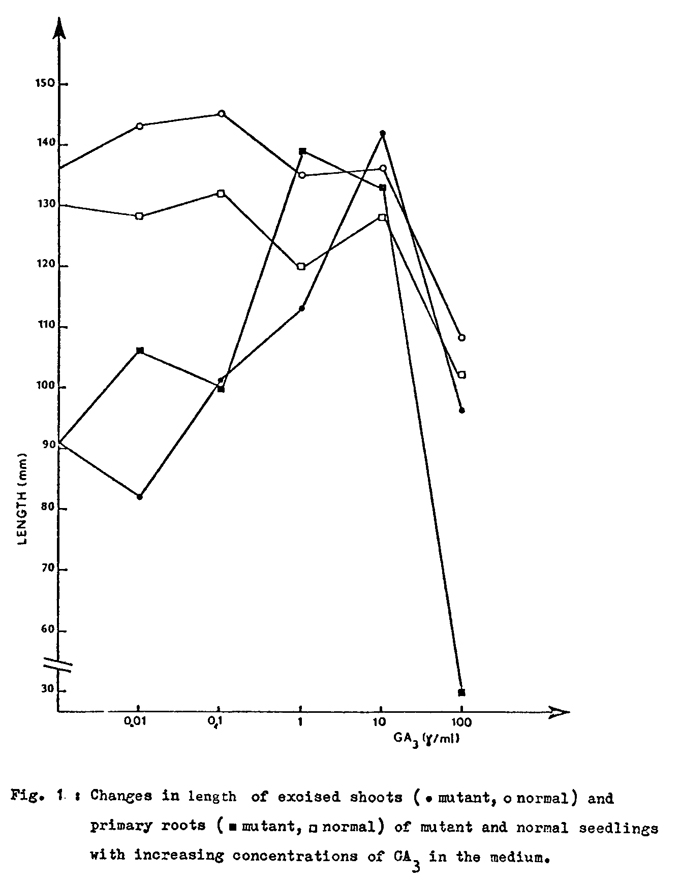
The data to be presented refer to the analysis of a series of seedling lethal mutants, isolated in the M2 generation following pollen mutagenesis (Neuffer, 1973). Only seven out of 50 mutants originally isolated were chosen for this test because of their property to express lethality at a very early stage of development. They were grown, as embryo cultures, on both a mineral (D) and an enriched (D+) medium (Gavazzi et al., 1975). Only one mutant, referred to as E283B, showed a slight improvement in its growth if cultured on D+ rather than D medium. This improvement becomes more evident if the mutant is allowed to grow for 24 rather than 12 days. Mutant E283B is easily distinguishable from normal siblings because of the olive pigmentation, necrosis of the leaf tissues starting before the first leaf emergence, and stunted growth. The latter feature might reflect an alteration in hormone metabolism. Accordingly the effect of IAA, GA3 and kinetin on mutant growth was tested by adding these hormones separately to a liquid mineral medium and allowing excised mutant shoots, 1 mm long, to grow on these media for 15 days. The results (Table 1) indicate that GA3 has a slight growth promoting effect on the mutant shoots. The effect of increasing doses of GA3 on root tips and shoots cultured on liquid media is given in Fig. 1. In both organs GA3 elicits a positive growth response. The best response is observed at 1 and 10 g/ml for roots and shoots respectively. At these concentrations the mutant appears to grow as much as the normal sibling. The effect of GA3 was also tested by incubating root and shoot tips on a mineral medium and then transferring them to a fresh medium supplemented with GA3 (10 g/ml). The aim of the incubation is to deplete the organs of their endogenous gibberellins. The results (Table 2) confirm the capacity of GA3 to induce growth of the mutant organs. The phenotypic repair applies to growth, but not to the "olive" phene. It looks as if a single mutational event has led to simultaneous loss of two metabolic functions, one resulting in growth impairment and the other affecting the photosynthetic pigments. Only the former function is restored by GA3 administration. To account for this observation one might hypothesize that the mutation E283B affected an early step of a branching metabolic chain lying before the branch point. In this case administration of GA3, one of the two end products of this pathway, would lead to repair of only one of the two missing functions. A branching point in the biosynthesis of gibberellins is in fact known and is represented by the synthesis of geranylgeranyl pyrophosphate, a common precursor of carotenoids, diterpenes, phytol and gibberellins. Further work with the olive necrotic mutant is aimed at proving whether in fact the above hypothesis can be validated.
Media composition: The H medium for root tip growth is the Heller medium, while that for shoots (F medium) is composed of major salts as in White (1943), minor elements as in Nitsch (1951) and vitamins as in White (1963). The composition of the D mineral and enriched medium for embryos culture is that already employed in previous tests (Gavazzi et al. MNL 47). (Research financially supported by NATO Research Grant No. 950).
M. Motta, G. Gavazzi, M. L. Racchi and M. G. Neuffer
Return to the MNL 51 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}