Alcohol dehydrogenase (ADH, E.C.1.1.1.1.) is an advantageous model for studying polyploidy effects at the molecular level. The maize tetraploid lines used in this analysis were obtained by the method of nonreduced gametes. The relative contributions of diploid lines to the genotypes of the tetraploids (RCD) were different (from 7/8 of the genotype to 15/16). The RCD depended on the number of backcrosses of the tetraploid to the initial diploid line. The following diploid-tetraploid pairs were analyzed: Gb627(2x)-Gb627(4x), Gb300(2x)-Gb3004x).
We also used diploid line R109 and line R109(4x), which was identical to this diploid line. R109(4x) was obtained by heat shock (L. F. Randolph, 1937).
Haploids were obtained by the method of Chase (Chase, 1947; Nanda, Chase, 1966) modified in our laboratory. The homozygous Adh-S Adh-S diploid line W155 was crossed to the homozygous Adh-F Adh-F marker line with purple endosperm and embryo. Haploid plants were identified by the absence of coloring in embryos at the seed stage, with subsequent electrophoretic analysis of ADH. Alcohol dehydrogenase activity was assayed in the scutellum of dry maize seeds.
The results of the comparisons of ADH activity in diploids and tetraploids of maize lines are given in Table 1.
We introduced index a = T-D/D to estimate quantitatively differences in the activities of this enzyme in diploids and tetraploids of the compared pair (T and D are the specific activities of the tetraploid and diploid). This index varies largely in different lines. The data obtained evidence that the degree of the differences in ADH activity between diploids and tetraploids is determined by genotypic environment.
Table 2 summarizes the results of the comparisons of ADH activity in haploid and diploid plants of line W155 and in plants of the marker line.
As Table 2 shows, ADH activity in maternal and paternal haploids is close to its activity in the diploid line W155 and significantly different from ADH activity in the marker line. This permits us to exclude, in this case, the maternal effect in ADH activity. Thus, there were no differences in ADH activity in line W155 diploids and haploids. This was attributed to either genotypic features of the line or to some specificities in the transition from the diploid to the haploid level.
Comparisons of ADH activity in diploids and tetraploids during growth demonstrated that differences in ADH activities between diploids and tetraploids are conditioned by the developmental stage and environmental conditions.
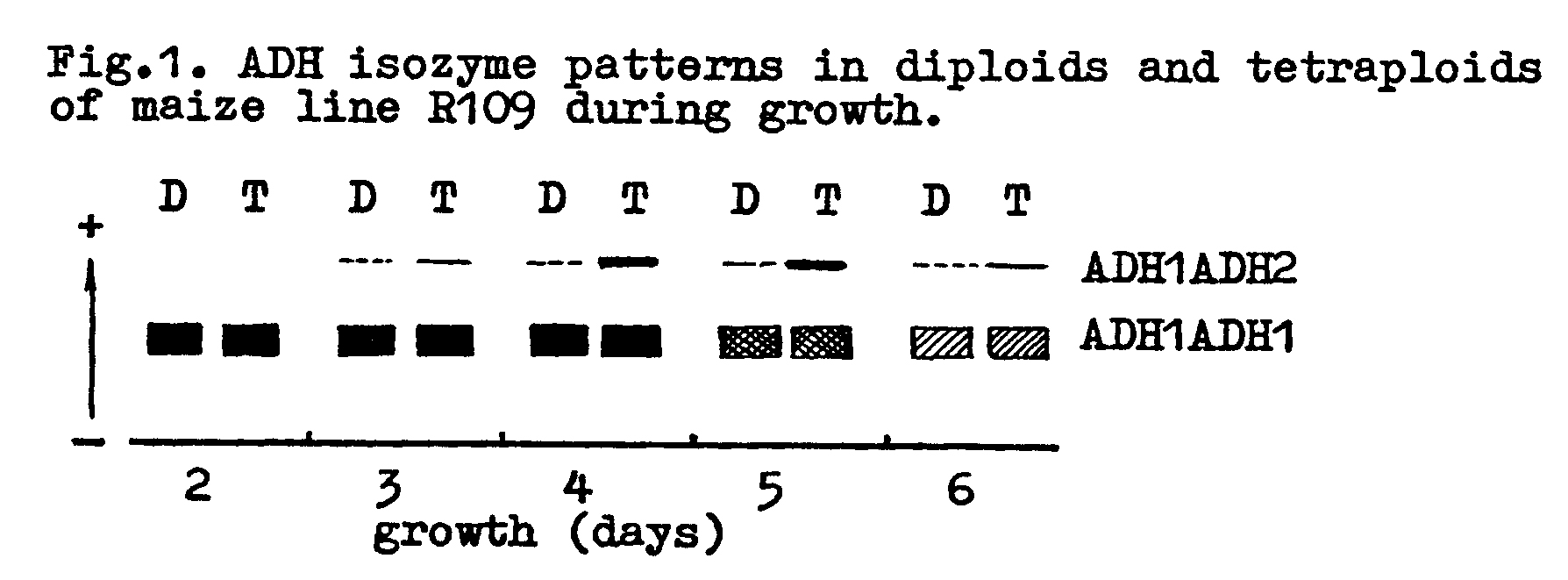
The relative expression of ADH at the structural loci was investigated, in view of the fact that ADH in maize is controlled at two non-linked loci, Adh and Adh2 (Schwartz, 1969; Freeling and Schwartz, 1973). The ADH isozyme patterns in diploids and tetraploids of line R109(2x) and R109(4x) during growth are represented schematically in Fig. 1.
The relative intensity of the heterodimeric zone in tetraploids is higher than in diploids at all growth stages. This indicates that changes in ploidy level are associated with changes in the relative expression of genes Adh and Adh2.
Thus, maize ADH is a good case illustrating that polyploidy changes ADH activity in maize tissues and modifies the phenotypic expression of genes controlling ADH. It may be suggested that polyploidy affects similarly the activity of many enzymes, which control biochemical processes underlying the formation of morphological and physiological characters in plants.
E. V. Levites and T. I. Novojilova
Return to the MNL 51 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}