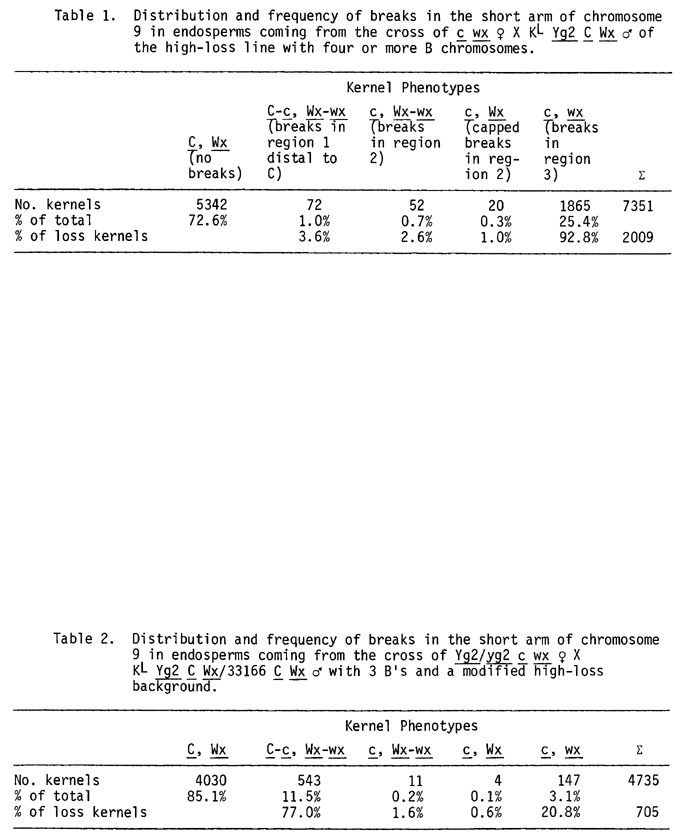
In the high-loss strain first studied by Rhoades, Dempsey, and Ghidoni in 1967, a knobbed chromosome 9 undergoes chromatin elimination in the second division of microspores with two or more B chromosomes. One of the two sperm has a normal chromosome and the other a deficient one. Fertilization of egg and polar nuclei by the dissimilar sperm appears to be random. Breaks at any position proximal to the Yg2 locus in the short arm of 9 produce a deficient chromosome which results in a yellow-green F1 seedling in testcrosses when the deficient sperm unites with the egg. When the deficient sperm with a freshly broken end fuses with the polar nuclei, break positions can be localized to specific regions in the arm. A break between C and the knob (region 1) gives a broken chromosome with the C and Wx alleles, which undergoes the bridge-breakage-fusion cycle in the endosperm mitoses resulting in kernels with variegation for C and Wx. Breaks between C and Wx (region 2) yield wholly colorless kernels which are variegated for Wx or wholly Wx when the broken end is capped, while breaks proximal to Wx (region 3) give colorless and waxy endosperms (Table 1).
Following a cross of yg2 c wx with pollen from a KL Yg2 C Wx high-loss plant, one of the seedlings from a colored kernel (no loss in the endosperm) exhibited the recessive yg2 phenotype--i.e., it had received a chromosome 9 deficient for the Yg2 locus from the pollen parent and a normal 9 with the yg2 allele from the egg parent. This particular plant (33166) had normal appearing pollen. When used in reciprocal testcrosses, 33166 gave approximate 1:1 ratios for C:c and for Wx:wx both as female and as male parent. Clearly, this chromosome 9 was deficient for Yg2 but had the C and Wx alleles, and the distal deficient segment carried no genes essential for male and female gametophyte development and functioning. Surprisingly, the C kernels from the male testcross included many which had a bridge-breakage-fusion type of variegation for the C and Wx loci. The loss phenomenon was not expected in this cross since the deficient 9 presumably had lost the terminal heterochromatic knob when the yg2 deficiency was produced. Cytological examination revealed that the 33166 chromosome was of the same length as a normal knobless 9 but possessed at the tip of its short arm a small heterochromatic knob--i.e., the 33166 chromosome, deficient for a short terminal piece including the Yg2 locus, had been capped by a small knob of uncertain origin. It may represent the small knob at the end of 7S since no chromosome of the high-loss line other than 9 and 7 had terminal knobs, but this surmise has not been verified. We consider it unlikely that the small knob on the 33166 chromosome is a remnant of the large knob on the parental chromosome 9 because such a constitution would require two closely spaced breaks at the tip of 9S accompanied by the deletion of the acentric fragment with most of the knob.
The possibility that the unexpected mosaic kernels were due to a modified type of chromatin elimination was investigated by comparing the behavior of a normal KL9 and the 33166 chromosome in heterozygotes with 1B or 3B's. Table 1 shows the loss rate in KL9/KL9 homozygotes from previous studies while Tables 2 and 3 give the data from male parents heterozygous for KL9 and 33166, having 3B's and 1B, respectively. In heterozygous plants, breaks at the second microspore mitosis may involve either the KL9 or the 33166 chromosome. Previous studies have shown that loss of KL9 occurs only in spores with two or more B's. Therefore, no loss of KL9 is expected in the 1B plants of Table 3 and a reduced loss frequency should be detected in the 3B plants of Table 2. Since over 90% of the breaks in the KL9 chromosome are proximal to the Wx locus in microspores of KL9 homozygotes of the high-loss line, it appeared likely that the minority class of c wx kernels of Table 2 came solely or chiefly from chromatin loss in 2B spore-s with the KL9 chromosome. This possibility was tested by determining the genotype of the embryos in C-c Wx-wx and in c wx kernels. If, for example, the c wx kernels all came from loss of the KL9 chromosome with the Yg2 allele, the ensuing seedlings (no loss in embryo) would be green while involvement of the Yg2-deficient 33166 chromosomes would yield green and yellow-green seedlings in a 1:1 ratio, due to heterozygosity for yg2 in the tester female parent. The tests made to date reveal that in microspores from KL9/33166 heterozygotes the vast majority of breaks occur in the 33166 chromosome and that the KL9 chromosome has a low rate of chromatin elimination in 2B microspores. This can be ascribed to the changed background of genetic modifiers in the KL9/33166 heterozygotes, relative to those in the original high-loss strain, which differentially affects the KL9 and the 33166 chromosome. Moreover, as is clearly shown in Table 3, the 33166 chromosome is capable of loss in spores with only 1B, unlike the KL9 chromosome. In high-loss strains, a normal chromosome 9 with a small terminal knob (KS9) undergoes a low percentage of chromatin elimination at the second microspore division. However, in heterozygotes for KL9 and 33166, the 33166 chromosome with a small knob comparable in size to Ks has a rate of loss much greater than that of the normal chromosome 9 with its large knob.
The KL normal 9 and the derived 33166 chromosome differ in the distribution of breaks in 9S, as is shown by a comparison of the data in Tables 1 and 2. In the high-loss strain homozygous for the KL9 chromosome, breaks between the C locus and the terminal knob at the second microspore mitosis are relatively infrequent (3.6% in Table 1). The vast majority of breaks occur in the proximal half of the arm to the right of the Wx locus. In contrast are the data on break position in Table 2 (where breaks are occurring primarily in the 33166 chromosome). 77.0% of the breaks are distal to C and only 20.8% take place in the proximal half of 9S. Clearly the position of the primary break at the second microspore division differs in the KL9 and 33166 chromosome and the susceptibility of different regions of 9S to breakage is a heritable trait.
Expression of the differential breakage pattern by which the two chromosomes differ is restricted to the second microspore division. For example, if a chromosome with the C allele and a freshly broken end is produced at the second microspore mitosis, fusion of the two broken ends takes place following replication. The ensuing chromatid type of bridge-breakage-fusion cycle in the developing endosperm results in a kernel mosaic for colored (C) and colorless (loss of C) sectors. The pattern of mosaicism is determined by the position of bridge breakage and there is a wide range in the size and distribution of colored spots between different mosaic kernels. However, the mosaic patterns amongst kernels arising from breaks distal to C in either the original high-loss chromosome 9 or in the 33166 chromosome are not sensibly different. In other words, although there is a striking difference between the two chromosomes in the position of breaks at the second spore mitosis, it is a difference which is not maintained during endosperm ontogeny.
No data are presently available for rate of chromatin loss in the microspores of plants homozygous for the 33166 chromosome since these homozygotes give lethal albino seedlings, as do wd/33166 heterozygotes. Viable (green-white striped) homozygotes are being synthesized by adding McClintock's Wd ring chromosome which possesses the Yg2 allele.
The data presented in this report suggest that a mechanism other than the postulated dicentric resulting from delayed knob replication in the second microspore division may be responsible for the inception of the bridge-breakage-fusion cycle. The nature of this mechanism can only be conjectured at this time, but it may be somewhat akin to the situation described by McClintock where breaks occur specifically at the Ds locus, which is transposable. There are significant differences between our high-loss phenomenon and Ac-Ds. In the former the position of the primary break, although occurring preferentially along the chromosome, is not restricted to one specific site as is true for Ds. Another difference is that the high-loss phenomenon involves the interaction of two known heterochromatic elements, B chromosomes and knobs, while the nature of Ac and Ds remains to be established. A third difference is the stage in development at which the primary breaks occur. In the high-loss phenomenon, the initial break takes place at one specific cell division while Ac-Ds breaks are not so restricted.
We have recently found a derivative of 33166 in which the majority of breaks take place between C and Wx rather than between Wx and the centromere for KL9 and between the knob and C for 33166. The response of this new chromosome to dosage of the B chromosome and any change in chromosome structure are being investigated.
M. M. Rhoades and Ellen Dempsey
Return to the MNL 51 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}