In the process of extending a gene dosage study of the Adh locus, it was found that the expression of certain enzymes is affected by aneuploidy in a precise manner. There is a negative correlation between the enzyme level and the number of long arms of chromosome one present. A discussion of this phenomenon and its implications is the subject of this and the following reports.
Those enzymes discussed in this report are alcohol dehydrogenase, ADH; glucose-6-phosphate dehydrogenase, G6PDH; 6-phosphogluconate dehydrogenase, 6PGDH; isocitrate dehydrogenase, IDH; and phosphoglucomutase, PGM. The Adh locus is carried in the portion of chromosome one that has been translocated to the centromeric portion of the B in the B-A translocation 1La. The Adh gene has been mapped to within 2 map units of lw in the long arm of chromosome one (Schwartz, 1971, Genetics 67:411). The locations of the other enzyme loci are unknown. The comparisons of enzyme activities of different levels of aneuploidy were always performed on kernels from the same ear. The tissue studied was the mature scutellum, separated from the endosperm.
The aneuploids were generated in the following way. A euploid female homozygous for one ADH electrophoretic variant was crossed by pollen from a hyperploid plant (1 B1B11B) that carried a different electrophoretic variant in the translocated arm. Such a cross will generate monosomics, disomics, and trisomics. The trisomics from this type of cross exhibit a small kernel phenotype and consequently were usually discarded. The remaining kernels are normal in size and consist of monosomic and disomic embryos. The monosomics can be distinguished from the disomics by excising a small portion of the scutellum and subjecting an extract of this to starch gel electrophoresis. The monosomic kernels have only a single electrophoretic variant, i.e. the female contribution, whereas the disomics have two electrophoretic variants present.
Trisomic vs. disomic comparisons were made on kernels produced by using a hyperploid female that had one electrophoretic ADH variant in the normal chromosome and a second variant in the two translocated arms, and crossing it by a diploid male, which carried a third electrophoretic variant of ADH. Kernels produced from this cross which had three electrophoretic variants present were considered to be trisomics. Kernels with two electrophoretic variants present in a 1:2:1 homo:hetero:homodimer ratio were considered to be disomic. Trisomic production was performed in this manner to minimize the difference in kernel size in the two classes. In some cases, trisomics were produced by crossing a hyperploid male having a different variant. Trisomic vs. disomic scutella, in this case, can be distinguished by isozyme band ratios.
Finally, tetrasomics were produced by using, as females, hyperploid plants that had one variant on the normal chromosome and a second on the two translocated chromosomes, in crosses with hyperploid males that were homozygous for a third electrophoretic variant. In crosses of this type, the entire series of 1-4 doses of the long arm of chromosome one can be distinguished. The class of kernels with only one electrophoretic variant may be slightly ambiguous. Most of the kernels of this type should be monosomics, although some disomics may be of this type due to crossing over between the centromere and the Adh locus so that both the normal and translocated chromosome introduced into the egg carry the same allele. If this egg is fertilized by a sperm deficient for the long arm of chromosome one, the resulting scutellum would be indistinguishable from a monosomic scutellum, which has only one ADH variant. However, controls show that recombination between the normal A chromosome and the two B-A chromosomes is minimal in the hyperploids of the type used. An example of a cross to produce tetrasomics is as follows. If a hyperploid, ADH CFF plant with the F allele on the translocated chromosome were used as a female, three major classes of gametes are formed: C, F, CF. If this plant is pollinated by a hyperploid male of ADH SSS constitution, each class of gamete could unite with a sperm that has no S, one S, or two S's. The major ADH genotypes from such a cross are as follows: C, F, CF, CS, FS, CFS, CSS, FSS, CFSS. Each of these can be distinguished from the others on the basis of the ADH variants present and their ratios.
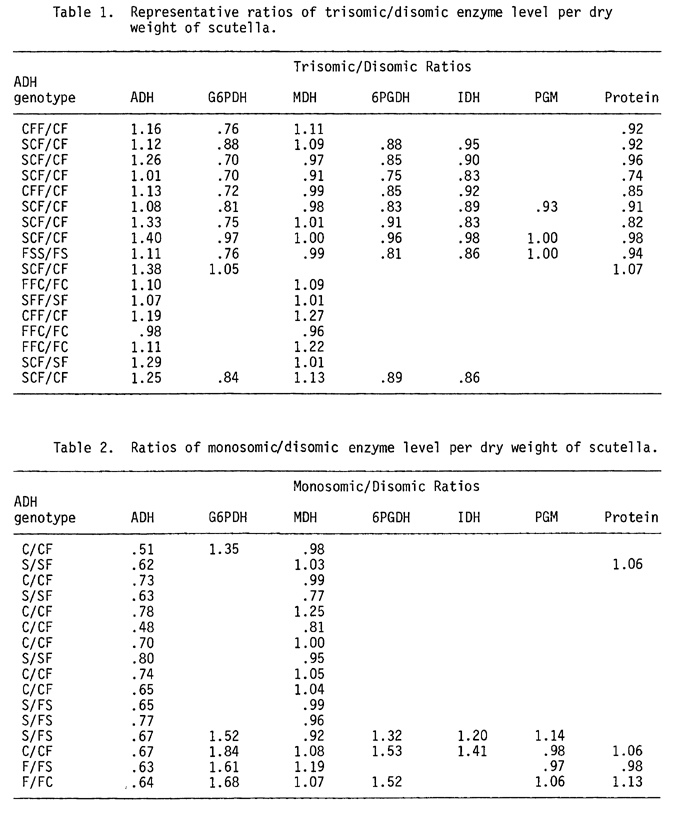
Representative trisomic/disomic enzyme ratios per dry weight of mealed scutellum are shown in Table 1. Monosomic/disomic enzyme ratios are shown in Table 2. The data in Tables 1 and 2 are from crosses, which are from diverse backgrounds. The variability among crosses was great and the values were not averaged. However, if one considers the modal values, certain trends become evident. From a strict dosage effect for Adh, one would expect the one dose/two dose ratio to be .50, but it is much greater than expected. Likewise, in three doses vs. two doses, the modal value is near 1.10 to 1.14. This value is much less than the 1.50 relationship expected from a dosage effect. The MDH and PGM values seem to be little affected by the aneuploidy. However, the G6PDH, 6PGDH and IDH values are altered. The G6PDH activity in trisomes is less than in disomes. In monosomes, the G6PDH activity is higher than in disomes. 6PGDH and IDH are not reduced to the same degree as G6PDH in trisomes nor are they increased as much in monosomics. Because of the values found for G6PDH, 6PGDH and IDH, it is doubtful that their structural gene loci reside in the long arm of chromosome one. This has not been rigorously proven, however.
Since some enzyme levels are affected but others apparently are not, the changes in enzyme expression are thought to be specific and not a general effect of aneuploidy on protein synthesis or metabolism. In order to test whether the change in enzyme levels associated with aneuploidy is the result of a change in the cell number per dry weight, DNA estimations were performed on scutella of trisomes, disomes, and monosomes. It was found that all three classes, after correction for the difference in chromosome number present per cell, had very similar amounts of DNA per dry weight. This shows that the aneuploidy in this case does not drastically change the number of cells per mass of tissue. The estimation of the hydrolysable DNA was by a modification of the method of Webb and Levy (J. Biol. Chem. 213:107-117). To test the sensitivity of the method, similar maize lines with 0 or about 8 B chromosomes were compared. The determined values were near those expected from given knowledge of the DNA content of a B chromosome (Ayonoadu and Rees, 1971, Heredity 27:365).
In order to test whether any qualitative differences in these enzymes were produced by aneuploidy, electrophoresis of extracts of mature scutellum was conducted and the gels were stained for the enzymes mentioned here by the methods (with modifications) of Brewer (An Introduction to Isozyme Techniques, 1970). MDH patterns were similar to those found by Yang and Scandalios (1974, Arch. Biochem. Biophys. 161:335). Zymograms of G6PDH, 6PGDH, IDH and PGM showed only a single major zone of activity. No discernible differences in isozyme pattern were seen between the aneuploid and euploids.
To test if there is any effect of B chromatin on the expression of these enzymes, other B-A translocations were used to generate aneuploids in which 1, 2 and 3 doses of the translocated portion could be phenotypically distinguished. Since six other aneuploid series do not give the same spectrum of enzyme expression as reported here for the TB1La series, it is concluded that the effects found are specifically produced by the A chromatin.
The tetrasomic analysis was conducted in a different manner due to the unavoidable circumstance that tetrasomic kernels are very small. The small size of the endosperm reduces the growth of the embryo. Since MDH activity did not seem to be affected by aneuploidy produced by TB1La in aneuploids of similar kernel size, ADH and G6PDH values were standardized against MDH. In one ear the standardized ADH trisomic/disomic and tetrasomic/disomic ratios were 1.19 and 1.17, respectively. G6PDH in the same ear gave ratios of 3/2 = .68; 4/2 = .54. In a second ear, ADH ratios were 1/2 .46; 3/2 = 1.19; 4/2 = 1.25. G6PDH values were 1/2 = 1.40; 3/2 = .83; 4/2 .67.
Considering all of the above data, it seems that the expression of G6PDH, 6PGDH and IDH is affected by a region of chromatin (a gene?) involved in TB1La in such a manner that there is a negative correlation between the dose of the varied region and the expression of these enzymes. In the case of G6PDH, the expression closely approaches the inverse of the ratio of the number of long arms of chromosomes one in the aneuploid to the number in the euploid. That is, in monosomics the ratio of expression of G6PDH approaches 2/1 (the inverse of 1/2) times the expression in disomics. In trisomics, the expression approaches 2/3 (the inverse of 3/2) the expression in disomics. For this reason the phenomenon has been tentatively referred to as the "inverse effect," and the chromosomal region responsible as the "inverse effect region" (IE region).
The equalization of ADH in these aneuploids deserves discussion. In this case the structural locus has been varied in a 1, 2, 3, 4 relationship, but the expression is not found to be directly proportional. One explanation for this could come from the gene competition hypothesis (Genetics 67:411). According to this interpretation, the expression is nearly equal in the aneuploid series because there is a limiting factor, produced elsewhere in the genome, that is required for ADH expression. Thus the ADH level would not depend on the number of structural genes present.
An alternative explanation is that a dosage effect of varying the Adh locus is cancelled by an inverse effect influencing ADH expression. Hence, when only one dose of Adh is present, there is also only one dose of an inverse region. Consequently, a reduction of ADH expression due to a reduced structural gene number is compensated by an increased expression due to the reduction in the number of IE regions. Any increase expected in the trisomic due to an increased number of structural genes would be cancelled by a reduction in expression due to the increased number of IE regions. Evidence that G6PDH, 6PGDH, IDH and ADH are all affected by the same IE region comes from exceptional ears in which ADH shows a dosage effect and G6PDH, 6PGDH, and IDH no longer show an inverse effect. This suggests, but does not prove, that the latter explanation for the compensation of ADH is correct. Experiments to distinguish these alternatives are in progress.
To understand the nature of the "inverse effect" more fully, these data from maize should not be considered alone. There is much evidence (although it has often gone unrecognized) for the presence of inverse regions in Drosophila and Datura (O'Brien and Gethman, 1973, Genetics 75:155; Rawls and Lucchesi, 1974, Genetical Research 24:59; Pipkin and Chakrabartty, 1975, Genetics 80:s64; Carlson, 1972, Molec. Gen. Genetics 114:273; Smith and Conklin, 1975, Isozymes, Vol. III). A discussion of their possible role in the regulation of gene expression, aneuploid depression, dosage compensation, sexual dimorphisms and sex determination is the subject of the other reports in this series. This report is only an abbreviated version of the data collected on this phenomenon.
James A. Birchler
Return to the MNL 51 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}