The effects of trisomy on kernel weight
Trisomy is being used as a tool to screen the maize genome for genetic loci that influence kernel weights. If a gene(s) influencing kernel weight and exhibiting gene dosage effects is present on a given chromosome, sibling kernels disomic and trisomic for this chromosome will have different kernel weights. By comparing the weights of sibling trisomic and disomic kernels, we are comparing the effects of three versus two copies of all genes on a specific chromosome.
The maize stocks employed in this study were generously provided by Dr. Gregory Doyle of the University of Missouri. Each trisomic type has been backcrossed three to eight generations to the inbred KYS to increase the isogenicity of each type. Thirty sibling kernels from the final backcross of each trisomic type were selected for analysis. Progeny kernels from the backcross to KYS of a female plant trisomic for chromosome 1 were designated family 1; kernels from the backcross of a female parent trisomic for chromosome 2 to KYS were designated family 2, etc. The smallest kernels from each trisomic type were selected for these experiments.
Each kernel employed in this study was dried in a Thelco model 18 forced-air oven at 40 C for 10 days then at 60 C for one week. These conditions reduce kernel water content to less than 1.3% w/w (Plewa and Weber, 1973. Can. J. Genet. Cytol. 15:313). This drying was necessary for nuclear magnetic resonance (NMR) spectroscopy studies that were carried out subsequently on the same kernels. The NMR studies will be discussed elsewhere. All kernels used in this experiment were dried simultaneously in the same oven; thus, the relative weights of the kernels should not be affected by this procedure.
After the kernels were dried, their weights were determined with a digital balance. The weight determinations were made prior to the identification of individual trisomic kernels; thus, possible experimental bias was absent from this experiment. To identify the trisomic kernels in each family, each kernel was germinated to produce a source of meristematic tissue for use in somatic chromosome counts. Root-tips were harvested from individual plants and were analyzed cytologically by the standard Feulgen squash technique. Due to rodent predation, the entire family segregating for trisomy of chromosome 9 was lost before the identification of trisomic kernels. Thus, no data are available on trisomic 9 kernels.
Plants that were identified as trisomics were transplanted to the field where they were allowed to grow to maturity. Approximately two weeks prior to anthesis, microsporocyte samples were removed from each plant and analyzed cytologically by the standard propiono-carmine smear technique. The identity of the trisomic chromosome was confirmed in aneuploid plants in each family.
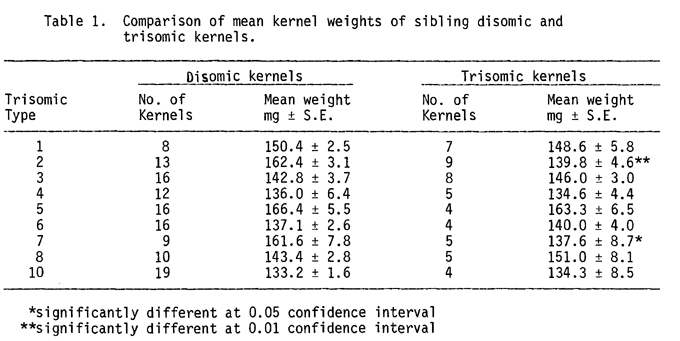
The results are presented in Table 1. The mean weight in mg of disomic kernels was compared with that for their sibling trisomic kernels using Student's t-test.
Because trisomy of specific chromosomes affects plant vigor in different ways, the progeny of distinct trisomic plant types will be affected differently by the specific aneuploid condition of the female parent. Variations in kernel weights between disomic kernels of different families reflect the distinct effects of trisomy of different chromosomes in their maternal parents. Hence, pooled data for all disomic kernels could not be compared accurately with pooled data for all trisomic kernels due to the considerable differences in kernel weights. However, the trisomic kernels in five of the families were lighter than their diploid sibling kernels, and in four families they were heavier. Thus, there was no general tendency for trisomic kernels to be either lighter or heavier than normal diploid kernels in this sample.
A comparison of kernel weights between sibling disomic and trisomic kernels within each family showed that significant differences existed in only two families. Trisomic 2 kernels were significantly lighter (p < 0.01) than their sibling disomic kernels. The mean weight for diploid kernels was 162.4 mg while the sibling trisomic 2 kernels had a mean weight of 139.8 mg. Trisomic 7 kernels were significantly (p < 0.05) lighter than their disomic sibling kernels. The mean kernel weights were 137.6 mg and 161.6 mg respectively. These data indicated that incompletely dominant genetic factors that influence kernel weight are present on chromosomes 2 and 7 of maize.
In family 8, the mean weights for sibling trisomic 8 and disomic kernels were 151.0 mg and 143.4 mg respectively. Although this difference is not significant at the 95% confidence interval, trisomic 8 kernels show a definite tendency to be heavier than their disomic siblings. Thus, incompletely dominant genetic factors that influence kernel weight may be present on chromosome 8 in addition to chromosomes 2 and 7 of maize.
Walter D. Fox and David F. Weber
Return to the MNL 51 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}