Maize Genetics Cooperation Newsletter vol 85 2011
CRA-MAC
Unit� di Ricerca per la
Maiscoltura
Via Stezzano,
24
24126 Bergamo (Italy)
USE OF BIOACTIVE PROTEINS IN PLANT
PROTECTION AGAINST PATHOGENS*
Lanzanova, C, Torri, A, Hartings, H, Berardo, N, Motto, M, Balconi, C
One of the main
topics of maize breeding is the improvement of plant protection against
pathogens. Plants respond to attack by pathogenic fungi with a complex network
of active responses
such as the production and accumulation of proteins that are
toxic or inhibitory to pathogens such as RIP (Ribosome Inactivating Protein).
The role of RIP in the pathogen defense has been documented (Balconi et al., 2010, Plant
Cell Monographs 18, Springer-Verlag Berlin Heidelberg : 149-166).
In maize
endosperm, a cytosolic albumin termed b-32 is synthesized
in temporal and quantitative coordination with the deposition of storage
proteins. In the past years b-32 was shown i) to enzymatically inactive ribosomes
modifying rRNA inhibiting protein synthesis in vitro (Maddaloni
et al., 1991, Journal of Genetics and Breeding, 45: 377-380); ii) to inhibit
the growth of Rhizoctonia solani
mycelia in an in vitro bioassay and plant assays (Maddaloni
et al., 1997, Transgenic Research, 6, 393-402). In this
context, we have recently shown and that maize b-32 was effective in wheat
transgenic lines as an anti fungal protein by reducing Fusarium culmorum head blight (FHB) (Balconi et al., 2007, European Journal of Plant
Pathology, 117, 129-140) and in maize transgenic lines reducing Fusarium verticillioides attack symptoms in leaf
tissues assays (Lanzanova et al., 2009, European Journal of Plant Pathology
, 124: 471-482).
Similarly to other RIPs, maize RIP is accumulated in
the seed as an inactive precursor, which is converted into an active form by proteolytic processing which removes peptide segments from
the N (residues 1-16 of pro-RIP) and C (residues 295-301) termini and also from
the center of the polypeptide (residues 162-186) (Hey et al., 1995, Plant
Physiol.107: 1323-1332).
Aims of the BIORES
project are devoted to deepen our knowledge about relationships between structure
and substrate specificity of b-32 protein, in order to clarify the role of the
processed segments of b-32 gene on
the ability of maize RIP to inhibit
fungal growth and/or mycotoxin accumulation.
Thereby, a series of
genetic constructions was made by selectively deleting the N-terminal, or
C-terminal or internal linker domain (Figure 1). Genetic deletions of the b-32 gene, that is apparently responsible for suppressing
enzymatic activity in the precursor, will be primarily expressed in Escherichia coli to produce sufficient quantities of modified
proteins.
To assess the role of
bioactive b-32 modified protein in the protection against fungal pathogens (F. verticillioides, Aspergillus flavus),
a series of in vitro bioassays are in
progress, to analyze their effect
on the fungal growth and on mycotoxin accumulation in comparison with i) the unmodified b-32 (control); ii) a commercial RIP (Saporin).
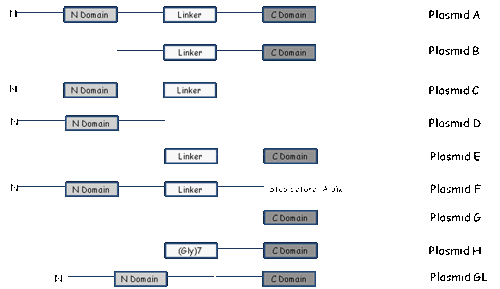
Figure 1: Genetic
deletions of the b-32 gene
Research
developed in the �BIORES- Use of
bioactive proteins in plant protection against pathogens - Utilizzo
di proteine bioattive nella protezione contro patogeni in pianta." CRA- Consiglio
per la Ricerca e Sperimentazione in Agricoltura-funded
project.
Please Note: Notes submitted to
the Maize Genetics Cooperation Newsletter may be cited only
with consent of authors.