Maize Genetics
Cooperation Newsletter vol 84 2010
Please Note: Notes submitted to the
Maize Genetics Cooperation Newsletter may be cited only with consent of
authors.
DNIEPROPETROVSK, UKRAINE
Institute of Grain Farming
Inheritance of
matroclinal haploidy in
diallel crosses
--Satarova, T N; Cherchel,
V Yu
Gynogenesis is an apomictic pathway of
reproduction when a new organism arises from an unpollinated egg cell and is
called a matroclinal haploid.
First S.S.Chase (Genetics 34: 328-332, 1949) has founded
matroclinal haploidy in maize. E.H.Coe (Am Nat 93:381-382, 1959) and
J.L.Kermicle (Sci 166:1422-1424, 1969) created marker
lines those permitted
taking as pollinators to identify haploids in populations and to stimulate to
some extent the ability to produce haploids. Now in maize breeding practice
matroclinal haploids after diploidyzation are widely used for rapid production
of homozygous lines.
The investigation of
genetic control of
matroclinal haploidy is actual both for the characterization of apomixis
in flowering plants and for the forecast of the structures of synthetic
populations which could become the initial material for selection of new
lines. Inheritable peculiarities
of this phenomenon should be taken into consideration also in programs of breeding lines with
high ability to haploid production to be used as testers in selection of marker
genotypes.
The aim
of the given investigation was to analyse the inheritance of the ability to
produce matroclinal haploids in maize. Genetic analysis was conducted in the
system of full diallel crosses according to B.I.Hayman (Biometrics 16: 369 - 381, 1960). Five lines were included to diallel
scheme, DК276-1, DК247, DК293, DК303/427, DК205/710,
and their reciprocal hybrids.
25 genotypes in total were organized in 4 repetitions. For production of matroclinal
haploids every genotype of
diallel scheme was pollinated with pollen of marker genotype Zarodyshevij
marker krasnodarskij 1 (ZMK-1). This marker had been selected by E.R.Zabirova et al.
(Kukuruza i sorgo 4: 17-19, 1996). It keeps dominant genes of plumule and endosperm coloration in purple. Seeds after
pollination with ZMK-1 of all the genotypes of diallel scheme were harvested
and haploids were identified
as caryopses with colourless embryos and coloured endosperm. Caryopses with
haploid embryos were sown next year into the soil and grown to anthesis to
verify their haploid status. �Frequency of matroclinal haploidy� was calculated
as the percentage ratio of the amount of caryopses with haploid embryos to the total amount of caryopses with
coloured endosperm. According to our observations after pollination of different
maize populations with pollen of ZMK-1 grain amount per ear is greatly declined
in comparison with free pollination (fig.1). Such a character was named
�degree of ear grainness�, calculated as the percentage ratio of average grain amount per ear after pollination with ZMK-1 to grain amount per ear of the same genotype
after free pollination, and also analysed in diallel crosses.

Figure 1.
Maize ears after pollination with marker genotype ZMK-1 show the declination of
grain amount.
The average values of
two traits for genotypes of diallel set are shown in table 1.
Table 1. The frequency of
matroclinal haploidy and the degree of ear grainness for inbred lines and
hybrids of diallel set.
|
Genotype |
Frequency of matroclinal haploidy,
%1 |
Degree of ear grainness, %1 |
|
DК 276-1 |
6,45�0,73 |
27,46�2,73 |
|
DК 247 |
7,79�1,23 |
22,93�4,14 |
|
DК 293 |
8,07�0,77 |
38,18�3,47 |
|
DК 303/427 |
5,62�0,71 |
42,60�4,14 |
|
DК 205/710 |
0,59�0,22 |
50,12�4,39 |
|
DК 276-1хDК293 |
5,05�0,77 |
17,94�2,79 |
|
DК 293хDК276-1 |
7,00�1,08 |
28,77�3,07 |
|
DК 276-1хDК247 |
6,27�1,03 |
22,86�3,61 |
|
DК 247хDК276-1 |
5,67�0,97 |
33,02�3,53 |
|
DК 276-1хDК303/427 |
5,73�1,04 |
28,71�3,95 |
|
DК 303/427хDК276-1 |
6,58�1,07 |
34,79�4,37 |
|
DК
276-1хDК205/710 |
2,36�0,63 |
13,01�2,63 |
|
DК
205/710хDК276-1 |
3,30�0,95 |
12,22�2,14 |
|
DК 293хDК303/427 |
7,42�0,76 |
47,35�7,16 |
|
DК 303/427хDК293 |
7,71�0,77 |
48,11�5,48 |
|
DК 247хDК293 |
10,68�0,89 |
33,26�4,94 |
|
DК 293хDК247 |
11,12�3,15 |
25,17�3,59 |
|
DК 293хDК205/710 |
6,42�0,92 |
26,13�6,45 |
|
DК 205/710хDК293 |
6,65�0,71 |
43,56�4,95 |
|
DК 247хDК303/427 |
7,89�0,78 |
36,98�4,89 |
|
DК 303/427хDК247 |
8,46�0,81 |
25,23�4,49 |
|
DК 247хDК205/710 |
5,31�0,65 |
37,64�4,83 |
|
DК 205/710хDК247 |
5,72�0,67 |
37,15�4,70 |
|
DК
303/427хDК205/710 |
4,51�0,61 |
42,19�4,43 |
|
DК
205/710хDК303/427 |
6,83�0,74 |
46,79�4,96 |
|
|
r* |
r0,95** |
|
For all
the experiment |
0,05 |
0,40 |
|
For hybrids |
0,33 |
0,44 |
|
For inbred lines |
-0,78 |
0,90 |
1-data are shown as X�mt0.05*
- Coefficient of pair correlation between frequency of matroclinal haploidy and
degree of ear grainness, ** - critical value of coefficient of pair correlation
at the 0,95 probability level.
In general frequency of matroclinal haploidy
in the diallel set fluctuated from 0,59% to 11,12%. Ear grainness after pollination with ZMK-1 compared to free pollination was reduced to 12,22-50,12%. It could be suggested that such
declination was connected
with the ability of genotype
to induce matroclinal haploidy.
However, in the given experiment coefficients of pair correlations between the
frequency of matroclinal haploidy and the degree of ear grainness in total for
the experiment, separately for hybrids and separately for lines were not significant. So, the
existence of such a relationship is not confirmed.
In the given diallel
set parental forms were presented by inbred lines, multiple allelism is
improbable, the data on the irregularity of meiotic chromosome disjunction are
absent. For the exclusion of
few differences between reciprocal hybrids their mean values were taken to
genetic analysis.
For �frequency of
matroclinal haploidy� the coefficient of regression Wr/Vr was
b=0,88�0,11 (tb=7,70, t1-b=1,03, t0,05=3,20), for �degree of ear grainness� it was b = 0,98 � 0,14 (tb
= 6,83; t1-b = 0,16; t0,05 = 3,20). The significance of regression
coefficients for both traits and their non-significant deviation from 1 testified
the absence of the effects of nonallelic interaction and dependent gene
distribution in parental forms. Therefore, additive and dominant genetic system
determines the manifestation
of two characters.
The analysis of variance of a
half of diallel table is represented in table 2.
Table 2. The analysis of variance
of �frequency of matroclinal
haploidy� and �degree of ear grainness� in diallel crosses.
|
Components of genetic variation |
Square sums |
Degrees of freedom |
Mean square |
F fact. |
F0,01 |
|
|
�Frequency of matroclinal haploidy� |
||||||
|
a |
0,6316 |
4 |
0,1579 |
65,93 |
4,22 |
|
|
b |
0,1690 |
10 |
0,0169 |
7,04 |
3,17 |
|
|
b1 |
0,0373 |
1 |
0,0373 |
15,56 |
7,82 |
|
|
b2 |
0,1011 |
4 |
0,0253 |
10,55 |
4,22 |
|
|
b3 |
0,0306 |
5 |
0,0061 |
2,55 |
3,90 |
|
|
R |
0,0094 |
3 |
0,0031 |
1,31 |
4,72 |
|
|
Rt |
0,0575 |
24 |
0,0024 |
|
|
|
|
�Degree of ear grainness� |
||||||
|
a |
1,2877 |
4 |
0,3219 |
19,05 |
4,22 |
|
|
b |
2,9134 |
10 |
0,2913 |
17,24 |
3,17 |
|
|
b1 |
1,5816 |
1 |
1,5816 |
93,58 |
7,82 |
|
|
b2 |
0,6934 |
4 |
0,1734 |
10,26 |
4,22 |
|
|
b3 |
0,6384 |
5 |
0,1276 |
7,55 |
3,90 |
|
|
R |
0,0453 |
3 |
0,0151 |
0,89 |
4,72 |
|
|
Rt |
0,4056 |
24 |
0,0169 |
|
|
|
For both traits the
significance of mean squares a and b testifies the effect of
additivity and dominance. The significance of b1 proves that the
effects of dominance are mainly concentrated in the same direction. The significance of b2 shows that
dominant alleles are not dispersed among lines identically. Mean square a includes not only
additive variance, but also a part of variance that is connected with dominant
effects. Mean square b3 is not significant for �frequency of
matroclinal haploidy� , so specific for every cross dominant effects which are
not connected with b1 and b2 are not established. Non-significant
R means weak effect of the
environment on the development of the characters.
On the diagram of
regression Wr/Vr for �frequency of matroclinal haploidy� the
regression line intersects the positive part of axis Wr (fig.2),
therefore, the middle degree of dominance for all the loci are incomplete, H1/D
< 1. Points of inbreds
DК276-1, DК247, DК293 and DК303/427 are located nearer to the start of
the regression line, they contain mainly dominant alleles (from 75% to 100%), which are
responsible for the decrease of �frequency of matroclinal haploidy�. For inbred
DК205/710 ratio
of dominant and recessive genes reaches the level 25% : 75%.
This inbred includes the biggest portion of recessive alleles, which
determine the increasing of matroclanal haploidy.
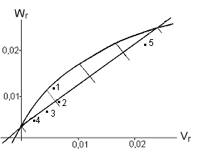
Fig.2. Regression ![]() for �frequency of matroclinal haploidy� in maize (1 – DК276-1, 2 – DК247, 3 – DК293, 4 – DК303/427, 5 – DК205/710).
for �frequency of matroclinal haploidy� in maize (1 – DК276-1, 2 – DК247, 3 – DК293, 4 – DК303/427, 5 – DК205/710).
For �degree of ear
grainness� the regression
line (fig.3) passes through the negative part of axis Wr, so the superdominance
plays the significant role in the manifestation of this character, H1/D > 1.
Dominant alleles are responsible for the decreasing, but recessive ones - for the increasing of ear
grainness. The distribution of dominant and recessive alleles
is located within interval 75%: 25% for inbreds DК293 and
DК303/427, for inbred DК276-1 it is near to 50%: 50%. For inbreds DК205/710 and
DК247 such
distribution approaches 25%:
75%.
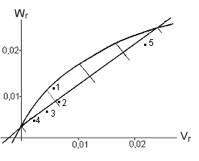
Fig.3. Regression ![]() for �degree of ear grainness� in maize (1 – DК276-1, 2 – DК247, 3 – DК293, 4 – DК303/427, 5 – DК205/710).
for �degree of ear grainness� in maize (1 – DК276-1, 2 – DК247, 3 – DК293, 4 – DК303/427, 5 – DК205/710).
The estimates of genetic parameters are represented in table 3.
Table 3. Genetic parameters for
�frequency of matroclinal haploidy�
and �degree of ear
grainness�.
|
Genetic parameters |
�Frequency of matroclinal haploidy� |
�Degree of ear grainness� |
|
D |
0,03�9,54Е-05 |
0,04�0,0005 |
|
H1 |
0,02�0,0003 |
0,23�0,0012 |
|
H2 |
0,01�0,0002 |
0,20�0,0011 |
|
F |
0,01�0,0002 |
0,02�0,0012 |
|
E |
0,002�3,89Е-0,5 |
0,02�0,0002 |
|
H1/D |
0,54 |
5,40 |
|
|
0,73 |
2,32 |
|
½ |
0,53 |
0,31 |
|
H2/4H1 |
0,18 |
0,21 |
|
Heritability in wide sense |
0,85 |
0,82 |
|
Heritability in narrow sense |
0,67 |
0,28 |
For
�frequency of matroclinal haploidy�
incomplete dominance takes place (H1/D=0,54), including separate loci (![]() =0,73). For �degree of ear grainness� superdominance is shown (H1/D=5,4), in that
number in every locus (
=0,73). For �degree of ear grainness� superdominance is shown (H1/D=5,4), in that
number in every locus (![]() =2,3).
=2,3).
For the
studied traits the estimate
of ½![]() differs from 1, the level of dominance varies in different loci. H1�H2,
so dominant and recessive alleles are spread irregularly among parental
inbreds. Positive estimate of F verifies the exceeding of the amount or effects
of dominant alleles over the
recessive ones in the given set of lines and hybrids.
differs from 1, the level of dominance varies in different loci. H1�H2,
so dominant and recessive alleles are spread irregularly among parental
inbreds. Positive estimate of F verifies the exceeding of the amount or effects
of dominant alleles over the
recessive ones in the given set of lines and hybrids.
For
the given characters the high values of heritability in wide sense proves the
primary effect of genotypic variance in the development of the characters. The significant value of heritability in narrow
sense for �frequency of matroclinal haploidy� (0,67) confirms the significant role of additive
gene effects and permits to make favourable prognosis in phenotype selection for high frequency of matroclinal
haploidy. It is important for
production of testers, which are necessary in breeding programs of new inducer genotypes. The definite role of
dominant effects in the development of
�degree of ear grainness�
is indicated also by the considerable difference
between the heritability in
wide sense (0,82) and the heritability in
narrow sense (0,28).