PIRACICABA, BRAZIL
Universidade de São Paulo
PONTA GROSSA, BRAZIL
Universidade Estadual de Ponta Grossa
Evidence of natural occurrence of paramutation at p1 locus
— Mondin, M; Gardingo, JR
The maize P-rr gene encodes a Myb transcription factor that activates the phlobaphene pigment pathway, regulating the flavonoid biosynthetic genes in kernel pericarp, cob, husk and skills (Chandler and Stam, Nature Reviews Genetics, 5:532–544, 2004; Sidorenko et al., Plant Mol. Biol., 39:11–19, 1999). To understand the transcriptional regulation of the P gene, promoter regulatory sequences of the allele P-rr, identified by Ac transposition, were tested in transgenic plants (Sidorenko et al., Plant J., 22: 471–482, 2000). Transgenic plant lines (P-ww) containing a GUS reporter gene and an enhancer fragment, called P1.2, produced F1 plants with reduced pigmentation on the pericarp when crossed with P-rr plants. The reduced pigmentation was transmitted to the next generations, being the original altered allele denominated as P-rr′ (Sidorenko et al., Plant Mol. Biol., 39:11–19, 1999; Plant J., 22:471–482, 2000; Sidorenko and Peterson, Plant Cell, 13:319–335, 2001). The paramutation was attributed to the P1.2 fragment, which probably altered the DNA methylation pattern and reduced the transcript levels of the P-rr allele (reviewed in Chandler and Stam, 2004). Natural occurrence of P paramutagenic alleles was recently described by Das and Messing (Genetics, 136: 1121–1141). However, the literature for description of P paramutagenic alleles is very poor.
In a program to study the inheritance of paramutagenic alleles and their effects during the inbreeding cycles, we have isolated a reduced pigmentation phenotype in the pericarp. Crosses between the Cusco, Piranão and ITA varieties activate an Ac transposon, producing a F1 generation expressing P-vv phenotype (Fig. 1). The F2 generation segregates and produces every possible phenotype related to the P locus as P-rr, P-ww, P-wr and several intermediate patterns of reduced pigmentation, as well as variegated pericarp. A series of cobs is presented in Figure 2. The variegated cobs always produce kernels with reduced pigmentation in the pericarp, and a gradient of color expression level can be seen. Some cobs presented a cream color that could not be related to any event or known allele, and was probably produced by new allele. On the other hand, some cobs presented a very intense full color in comparison to the original P-rr or P-wr. The increase on the level of color could be attributed to intensifier genes. We do not know about the occurrence of these genes in our material, but this possibility should be considered.
The kernel analysis was very difficult because the varieties involved present the alleles required to the anthocyanin pathway. Paramutation in the r locus could be seen in some ears, deduced from the occurrence of white pericap with color only at the top of the kernel (Fig. 3). Some kernels presented both paramutation events, and it should be interesting to investigate the interaction between these events in the same line.
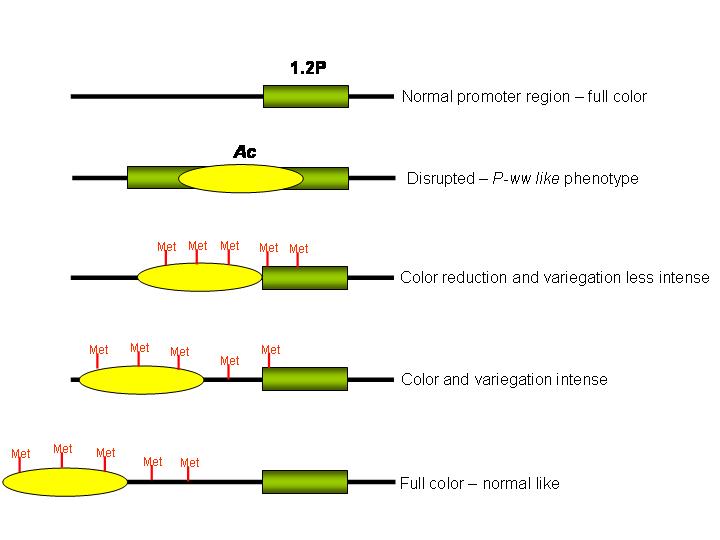
As cited in the review above, the level of color is strongly correlated to the increase of methylation and decreased transcript levels, and the paramutation events have been attributed to the 1.2P promoter region. Our idea is that the Ac transposable element could be inserted near the 1.2P promoter region (described earlier by Sidorenko et al.), altering the pattern of methylation and promoting the paramutation in our material. Probably, an insertion in the promoter region could generate a P-ww-like phenotype derived from a full-imprinting, whereas insertion near the promoter region could affect the transcription levels by alteration in the methylation pattern as a result of the transposable element inactivation. The effect of the paramutation would be the more intense the shorter the distance between the promoter region and the insertion (Fig. 4). However, our main objective is isolating the alleles and studying their behavior in crosses with other stable alleles. Thus, inbred lines of each derived paramutant phenotype have been produced.
Testing crosses have been conducted among those paramutant phenotypes and stable ones, well as among the P-vv and stable ones, to reproduce the phenomenon. We believe our hypothesis could be tested by mapping the position of the insertions in this way.
Figure 1: F1 P-vv phenotype produced after the crosses among the three varieties.
Figure 2: A series of ears where the arrows indicate the reduced color expression, possibly a P′ (paramutable) allele. Note a variation in the expression level; the two first (top row, from the left to right) ears show variegation and strong colored sectors, while the other ears show sectors with a reduced color expression.
Figure 3: Segregating ears showing paramutation phenotypes in the pericarp. The arrows indicate kernels that contain paramutation events in the locus p and r. These kernels have been isolated to study the interaction of different events of paramutation in the same plant. The other three ears (at right) show the difficulty in screening the pericarp paramutants with other loci segregating for kernel color.
Figure 4: Hypothetical model showing how the Ac insertion, its inactivation by methylation, and the distance of promoter region in the p locus would affect the paramutation.
Return to the MNL Volume 79 Index
Return to the index of Maize Newsletters
Return to the Maize Genome Database Page
{kind=link}
{kind=link}
{kind=link}
{kind=link}