BUENOS AIRES, ARGENTINA
Universidad Nacional de La Plata
BUENOS AIRES, ARGENTINA
Universidad Nacional de Lomas de Zamora
Meiotic pairing in the hybrid (Zea mays × Zea diploperennis) × Zea luxurians
— del Carmen, MM; Chorzempa, SE; García, MD
Genomic formulae, chromosome pairing, and the cryptic intergenomic pairing (induced by using a diluted colchicine solution) were analyzed in the tri-hybrid (MDL), obtained by crossing the hybrid MD (Z. mays cv. Colorado Klein × Zea diploperennis, both parents 2n=20) with Zea luxurians (2n=20).
Immature tassels were fixed in Farmer solution (3:1 absolute ethanol-acetic acid) and colored with a 2% solution of acetic haematoxylin (Nuñez, 1968). Pairing configurations were observed at diakinesis-metaphase I. Some of the immature tassels were treated with a 0.5 × 10-4 M colchicine solution during 12 h and washed in tap water during 24 h (Poggio et al., 1990), and finally control and treated plants were fixed in Farmer solution.
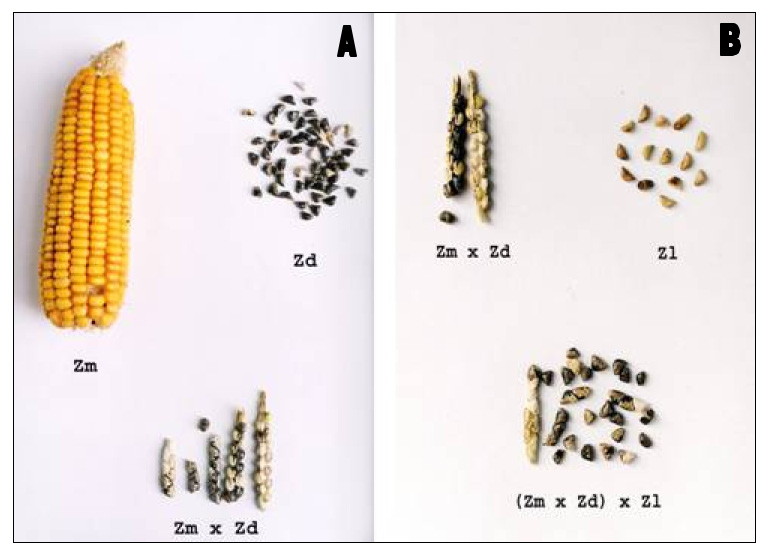
MDL plants were perennial and produced very prolific tillers and viable seeds. They showed a qualitative short day response of flowering to photoperiod. Caryopses were enclosed in cupules similar to those of teosintes and disposed on a rachis, which disarticulates at maturity in a similar way as wild Zea species do (Figure 1 A, B). Shoots arose from tassels (Figure 2 A, B) under specific environmental conditions such as high temperatures and long photoperiods. Plants can be obtained from these shoots by rooting them in water or moistened sand (Figure 2 C, D).
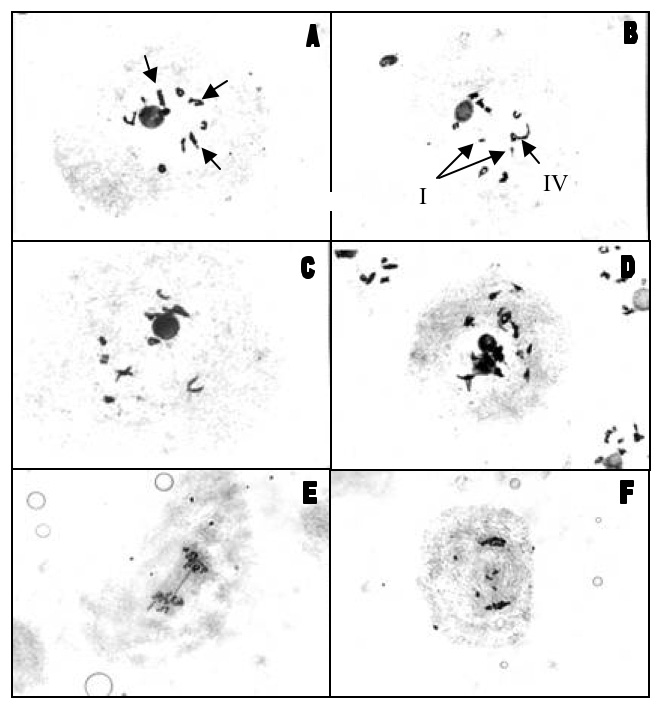
Cytogenetic analysis (Table 1) revealed an average number of uni-, bi- and tetravalents of 0.783I + 9.544II + 0.024IV. The most common meiotic configuration was 10II, with an average number of 14.57 chiasmata cell-1. Low frequencies of uni- and tetravalents (Figure 3 B) or cells with duplicated chromosome numbers were also observed (Figure 3 D). Bivalents were the result of chromosome pairing from Z. mays or Z. diploperennis with Z. luxurians. Three of the bivalents showed pairing between chromosomes of different length (Figure 3 A). The longest ones were Z. luxurians chromosomes. Cells showed a normal anaphase I. Only exceptionally, one bridge (Figure 3 E), lagging chromosomes (Figure 3 F) or cells with different chromosome numbers were observed.
In order to analyze the cryptic pairing between homoeologous Zea chromosomes, MDL tassels were treated with diluted colchicine solutions. Treated hybrid plants showed no significant differences with control plants. Similar results were observed in Z. diploperennis.
According to these results and previous ones related to evolutive cytogenetic of genus Zea (Molina and Naranjo, 1987; Naranjo et al., 1994, Molina and García 1999, 2001, 2004; among others), it can be suggested that chromosomes of the three species are homoeologous, and the MDL tri-hybrid genomic formulae is: 5II (Am or Ad) Al + 5II (Bm or Bd) Bl, where Am, Bm, Ad, Bd, Al and Bl are genomes A or B of Z. mays, Z. diploperennis and Z. luxurians, respectively.
Acknowledgements
This work was supported by CONICET, PIP 4650: Universidad Nacional de la Plata and Universidad Nacional de Lomas de Zamora.
References cited
Molina M. del C. and Naranjo C. A., (1987). Cytogenetic studies in the genus Zea. I. Evidence for five as the basic chromosomes number. Theor. Appl. Genet. 73: 542–550.
Molina M. del C. and García M. D., (1999). Influence of ploidy levels on phenotypic and cytogenetic traits in maize and Zea perennis hybrids. Cytologia 64: 101–109.
Molina M. del C. and García M. D., (2001). Ploidy levels affect phenotype and cytogenetic traits in Zea mays ssp. mays (2n=20 or 40) and Zea mays ssp. parviglumis hybrids. Cytologia 66: 189–196.
Molina M. del C., García M. D., López, C. G., Moreno Ferrero, V., (2004). Meiotic pairing in the hybrid (Zea diploperennis × Zea perennis) × Zea mays and its reciprocal. 141(2): 135–141.
Naranjo C. A, Poggio L., Molina M. del C. et al., (1994). Increase in multivalent frequency in F1 hybrids of Zea diploperennis × Zea perennis by colchicine treatment. Hereditas 120: 241–244.
Nuñez O., (1968). An acetic haematoxylin squash method for small chromosomes. Caryologia 21: 115–119.
Poggio L., Molina M. del C. and Naranjo C. A., (1990). Cytogenetic studies in the genus Zea. 2 Colchicine-induced multivalents. Theor. Appl. Genet. 79: 461–464.
Table 1. Average number uni-, bi-, tri- y tetra- per pollen mother cell (PMC) in species and hybrids of Zea. Numbers between parentheses represent minimum and maximum absolute values for each group.
| Species and hybrids | 2n | Meiotic configuration | Chiasmata Cell-1 | No. of PMC | |||
| I | II | III | IV | ||||
| Zd | 20 | 0.27 (0-4) | 9.73 (8-10) | 0 (0) | 0 (0) | 14.00 | 100 |
| Zm | 20 | 0 (0) | 10 (10) | 0 (0) | 0 (0) | 15.76 | 200 |
| Zl | 20 | 0.20 (0-4) | 9.80 (8-10) | 0 (0) | 0 (0) | 14.59 | 176 |
| MD | 20 | 0.85 (0-6) | 9.25 (7-10) | 0 (0) | 0 (0) | 15.00 | 181 |
| MDL | 20 | 0.78 (0-2) | 9.54 (4-20) | 0 (0) | 0.05 (0-1) | 14.57 | 166 |
Figure 1. Zea species and hybrids: A- Ears of Z. mays, Z. diploperennis and its hybrid; B- Female inflorescences of the hybrid Z. mays × Z. diploperennis, Z. luxurians and the tri-hybrid (Z. mays × Z. diploperennis) × Z. luxurians.
Figure 2. Tri-hybrid (Z. mays × Z. diploperennis) × Z. luxurians: A- Plants showing shoots originated from the tassel; B- Tassel showing development of shoots; C- Rooting shoots excised from the tassel; D-Plants regenerated from the tassel.
Figure 3. Meiotic configurations of the tri-hybrid (Z. mays × Z. diploperennis) × Z. luxurians: A- 10 bivalents. Arrows point different length paired chromosomes; B- One tetravalent and two univalent (arrows); C- 10II disposed in two groups of 5 each; D- Cell with duplicated chromosome number; E- Anaphase showing one bridge; F- Anaphase showing lagging chromosomes.
Return to the MNL Volume 79 Index
Return to the index of Maize Newsletters
Return to the Maize Genome Database Page
{kind=link}
{kind=link}
{kind=link}