IRKUTSK, RUSSIA
Siberian Institute of Plant Physiology and Biochemistry
An influence of cold stress on different mitochondrial uncoupling systems function of maize shoots
— Grabelnych, OI; Kolesnichenko, AV; Pobezhimova, TP; Koroleva, NA; Korzun, AM; Voinikov, VK
Previously it was shown that under cold shock (0 and -4°C, 1 h) living maize shoots can generate the heat for the initial 20 min (Kolesnichenko et al., MNL, 75:22, 2001). Recently it was found that there are at least three thermogenic systems in winter wheat shoots, connected with function of alternative cyanide-resistant oxidase (AOX), free fatty acids and UCP-like uncoupling proteins, and stress protein CSP 310 (Grabelnych et al., J. Therm. Biol., 28:571–580, 2003). It is known that AOX (Stewart et al., Plant Physiol., 92:761–766, 1990), UCP-like plant uncoupling mitochondrial protein (Jezek et al., J. Bioenerg. Biomembr., 32:549–561, 2001) and CSP 310 and CSP 310-like proteins (Kolesnichenko et al., J. Therm. Biol., 25:245–249, 2000) are also present in maize mitochondria protein spectra, but their thermogenic effect during cold stress in maize shoots was not studied previously. So, the aim of the present work is to study the heat generation by different thermogenic systems (AOX, UCP-like proteins and CSP 310) in maize seedlings shoots during cold stress.
The measurement of the temperature of chilling samples was performed as described previously (Vojnikov et al., Biochem. Physiol. Pflanzen. 179:327–330, 1984) by a copper-constantan thermocouple (wire diameter 0.1 mm) connected to the input of a high-sensitive microvoltmeter. The sensitivity of this thermocouple was 0.025°C.
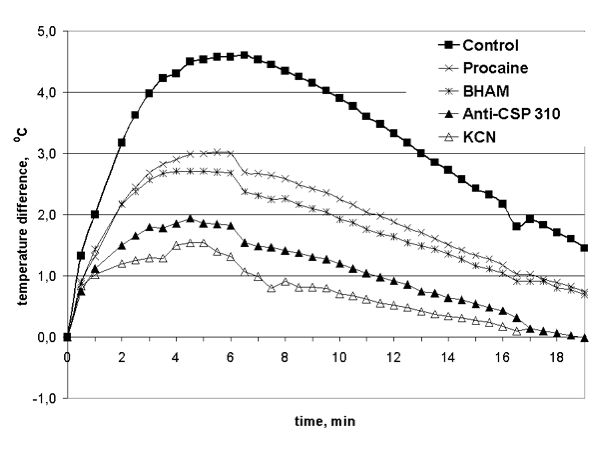
When maize seedlings were exposed to cold shock at -6°C, the temperature difference between “alive” and “killed” seedlings shoots (“Control”) was up to 4.0–4.6°C during the 4.5–7.5 min of cold shock (Fig. 1). Further chilling of maize shoots caused the reducing of temperature difference between them. It was found, that infiltration of shoots by KCN, that inhibit the transport of electrons on the main path of respiratory chain and keep function only AOX, caused the decrease of the maximum of temperature difference between “KCN treated alive” and “killed” maize shoots at the first 5 min of a cold shock from 4.5 to 1.5°C (Fig. 1). The infiltration of a winter wheat shoots by BHAM that inhibits the transport of electrons through the AOX caused the decrease of a temperature difference between “BHAM treated alive” and “killed” maize shoots at the first 5 min of a cold shock from 4.5 to 2.7°C (Fig. 1). The infiltration of a winter wheat shoots by procaine that inhibits phospholipase A2 activity and decrease the quantity of free fatty acids, which are substrates of UCP-like plant uncoupling mitochondrial proteins, as well as infiltration of shoots by BHAM reduced heat generation of maize shoots. The decrease of a temperature difference between “procaine infiltrated alive” and “killed” shoots at first 5 min of cold shock from 4.5 to 3.0°C was marked in this case (Fig. 1). The infiltration of maize shoots by antiserum against stress protein CSP 310 caused the decrease of the difference between “anti-CSP 310 infiltrated alive” and “killed” winter wheat shoots during first 5 min of cold shock from 4.5 to 2.0°C (Fig. 1). So, based on the data obtained we can conclude that three thermogenic systems function in maize during low-temperature stress.
Acknowlegements: The work has been performed, in part, with the support of the Russian Science Support Foundation, Russian Foundation of Basic Research (project 03-04-48151 and 05-04-97231).
Fig. 1. Temperature difference between water infiltrated and “killed” (“Control”), procaine infiltrated and “killed” (“Procaine”), BHAM infiltrated and “killed” (“BHAM”), anti-CSP 310 antiserum infiltrated and “killed” (“Anti-CSP 310”) and KCN infiltrated and “killed” (“KCN”) maize shoots at -6°C.
Return to the MNL Volume 79 Index
Return to the index of Maize Newsletters
Return to the Maize Genome Database Page
{kind=link}