BELGRADE, YUGOSLAVIA
Maize Research Institute
Changes in genetic gain from selection by the introduction of exotic germplasm into local synthetic populations of maize (Zea mays L.)
— Stankovic, G; Delic, N; Stankovic, S; Babic, M; Andjelkovic, V
INTRODUCTION
A long-term cultivation of maize hybrids as a practical application of heterosis has resulted in narrowing of genetic variability of the whole global maize population. According to many authors (2, 10, 19, 21), hybrid combinations grown in the USA and the southern parts of Europe have mainly been developed from the Iowa Stiff Stalk Synthetic. The other parental component in hybrids of that time was usually of the Lancaster base. It is believed that such a situation has remained unaffected. No more than a few populations (Reid Yellow Dent, Lancaster Sure Crop, Krug, Midland) have been maintained as a source of new inbred lines, for which it was considered that the loss of their genetic variability was about 30%.
The following methods are used with the purpose of increasing genetic variability of the breeding material: hybridisation of adapted material, mutagenic treatments, genetic engineering and introduction of exotic germplasm. There are different interpretations of the term “exotic germplasm of maize”. Some authors use the term “introduced maize” (18); others (16) consider maize from different regions exotic germplasm. The explanation that exotic germplasm is a source of germplasm which is not immediately applicable or adapted for certain breeding programmes (12) is perhaps the most common (i.e., exotic plasma is germplasm that encompasses new, useful alleles not present in adapted, local plasma (9)). Exotic germplasm can increase genetic variability of the adapted material, while recurrent selection is an efficient method for the further improvement of such material (12).
Genetic gain from selection depends on existing population variability, heritability level of the observed traits, selection intensity and the number of generations necessary for a cycle of recurrent selection (6). The grain yield increase of 18% per cycle was determined by the application of half-sib recurrent selection with the use of a single cross hybrid as a tester to a composite population of a broad genetic base (15).
MATERIALS AND METHODS
The material observed in this study was developed via three cycles of selection. The aim of selection was to improve local commercial inbred lines of the first cycle of Yugoslav selection (V395, V390, R348, Š144, R59) with the US inbred B37 (of BSSS background). The stated inbred lines were crossed to the inbred B37 and selfed progenies were developed. After 3–4 years of investigation, the best inbred lines were selected and used to develop the synthetic ZP/USA (local and introduced germplasm). The second synthetic, ZP, was developed by diallel crossings of stated inbred lines that had been derived from five synthetics by crosses of Istrian Gourd selected dents, Vukovarski dent, Šidski dent and Rumski dent (local germplasm). Self-pollination of randomly selected plants of both synthetics was performed with simultaneous crossing to a tester, the inbred line L75, which was used as a female component.
The material used in the trials consisted of 96 S1 families of the population Syn ZP, 68 S1 families of the population Syn ZP/USA, 96 HS (half-sib) families of the population Syn ZP and 68 HS (half-sib) families of the population Syn ZP/USA. Four separate trials were set up according to the nested design (incomplete block design with replications within a set (5)) and were carried out in four locations. The observed progenies were evenly distributed over four sets with two replicates within a set and 20 plants per replicate. Standard maize-growing practices were applied. The following traits were measured: 14% moisture grain yield (t ha-1), plant height (cm), ear height (cm), number of ears per plant, number of lodged and broken plants (%) and grain moisture content (%).
Based on mean square values obtained by the analysis of variance, the following genetic parameters were estimated:
Genetic variance (σ2g)
Standard error of genetic variance (SEσ2g)
Broad-sense heritability h2=(σ2g / σ2f ) × 100
Standard error of heritability (SE h2)
Expected genetic gain from selection ΔG=(c × k × σ2g) / (y × σf) (20).
RESULTS AND DISCUSSION
It is evident that the introduction of introduced germplasm did not affect grain yield increase in neither type of progenies of the population ZP/USA in relation to the population ZP (Table 1). According to some authors, hybrids developed from crossings of inbred lines that partially or completely contain exotic germplasm, overyield hybrids derived from adapted sources (1 and 11). On the other hand, other authors report that heterosis does not always occur in crossings of the adapted to the non-adapted material (8 and 16). Contrary to the grain yield reduction, the introduction of introduced germplasm into the local material resulted in improved means of the grain moisture content, stalk resistance to lodging and ear height. Considering that these traits are perhaps the greatest disadvantages of local materials, a certain positive advancement was made by the introduction of introduced germplasm.
Table 1. Means of studied traits in synthetic populations
| Trait | Syn ZP HS | Syn ZP S1 | Syn ZP/USA HS | Syn ZP/USA S1 |
| Yield (t/ha) | 9.10 | 4.40 | 8.70 | 4.20 |
| Plant height (cm) | 217.80 | 176.90 | 229.00 | 185.40 |
| Ear height (cm) | 96.90 | 72.70 | 99.20 | 72.60 |
| Ear per plant | 1.23 | 1.05 | 1.21 | 0.99 |
| Moisture (%) | 22.58 | 25.65 | 22.53 | 24.28 |
| B/L plant (%) | 5.90 | 15.40 | 8.80 | 11.60 |
According to results obtained in our studies, the values of genetic variance for grain yield in HS progenies of observed populations were not significantly different, while the difference in S1 progenies was highly significantly higher in the synthetic ZP/USA. The increase of genetic variance in populations per se that contain exotic germplasm is in accordance with the results obtained by other authors (4 and 17). Genetic variance for the grain moisture content is significantly increased in the population ZP/USA-S1 in relation to the population ZP-S1, while the same parameters were decreased for some observed traits (plant height, ear height, stalk resistance to lodging) or were insignificantly increased (number of ears per plant).
Table 2. Genetic variance (σ2g) and broad-sense heritability (h2) of studied traits in synthetic populations
| Traits | Syn ZP HS | Syn ZP/USA HS | Syn ZP S1 | Syn ZP/USA S1 | |
| Yield (t/ha) | σ2g(SE σ2g) | 0.47 *(0.17) | 0.53 *(0.22) | 0.37 *(0.10) | 1.27 *(0.27) |
| h2(SE h2) | 41.44(0.15) | 43.68(0.18) | 52.05(0.15) | 64.35(0.17) | |
| Plant Height (cm) | σ2g(SE σ2g) | 9.23 *(3.66) | 13.72 *(6.47) | 42.29 *(14.34) | 23.38 *(8.52) |
| h2(SE h2) | 38.97(0.15) | 39.14(0.18) | 45.15(0.15) | 49.77(0.18) | |
| Ear Height (cm) | σ2g(SE σ2g) | 4.69 *(0.002) | 6.82 *(2.99) | 17.47 *(4.28) | 15.29 *(4.36) |
| h2(SE h2) | 34.73(0.15) | 41.91(0.18) | 61.06(0.15) | 62.47(0.17) | |
| Ear per plant | σ2g(SE σ2g) | 0.00082 *(0.00031) | 0.00150 *(0.00077) | 0.00270 *(0.00075) | 0.00286 *(0.00092) |
| h2(SE h2) | 41.10(0.15) | 38.25(0.18) | 54.56(0.15) | 55.73(0.18) | |
| Moisture (%) | σ2g(SE σ2g) | 0.22 *(0.08) | 0.29 *(0.13) | 0.48 *(0.14) | 0.76 *(0.20) |
| h2(SE h2) | 40.02(0.15) | 40.40(0.18) | 52.98(0.15) | 54.91(0.18) | |
| B/L Plant (%) | σ2g(SE σ2g) | 56.51*(14.92) | 27.53*(11.23) | 85.69 *(24.93) | 24.19*(11.15) |
| h2(SE h2) | 56.67(0.15) | 44.79(0.18) | 52.03(0.15) | 40.01(0.18) | |
Significance: *(P<0.05), **(P<0.01), n.s.—non significant
Two populations (adapted germplasm and adapted introduced germplasm) were used in our studies to predict the improvement of grain yield and five observed traits on the basis of evaluation of S1 and HS progenies at selection intensities of 5%, 10% and 20% (Table 3). The highest expected genetic gain from selection of grain yield was estimated in the population ZP/USA-S1 and ranged from 11.14% to 16.42% at the selection intensities of 20% and 5%, respectively. These results are in accordance with other authors’ results (3 and 14). The application of recurrent selection resulted in grain yield which was higher by 2–4% per cycle in observed populations (13). The same author suggested that the selection intensity should have been lower (20%) in the initial cycles in order to avoid genetic drift.
Table 3. Expected genetic gain from selection (ΔG) for studied traits in synthetic populations at three levels of selection intensity (5%, 10%, 20%)
| Intensity 5 % | Syn ZP HS | Syn ZP/USA HS | Syn ZP S1 | Syn ZP/USA S1 | ||||
| Trait | ΔG | %* | ΔG | %* | ΔG | %* | ΔG | %* |
| Yield | 0.273 | 3.01 | 0.313 | 3.61 | 0.299 | 6.74 | 0.695 | 16.42 |
| Pl. height | 1.257 | 0.58 | 1.528 | 0.67 | 2.445 | 1.38 | 2.048 | 1.10 |
| Ear height | 0.810 | 0.84 | 1.089 | 1.10 | 2.105 | 2.89 | 2.029 | 2.79 |
| Ear/plant | 0.013 | 1.01 | 0.015 | 1.25 | 0.026 | 2.48 | 0.026 | 2.59 |
| Moisture | 0.169 | 0.75 | 0.198 | 0.88 | 0.299 | 1.17 | 0.418 | 1.72 |
| B/L plant | 3.955 | 66.44 | 2.330 | 26.47 | 4.384 | 28.34 | 1.765 | 15.20 |
| Intensity 10 % | Syn ZP HS | Syn ZP/USA HS | Syn ZP S1 | Syn ZP/USA S1 | ||||
| Trait | ΔG | %* | ΔG | %* | ΔG | %* | ΔG | %* |
| Yield | 0.233 | 2.56 | 0.266 | 3.07 | 0.254 | 5.73 | 0.592 | 13.97 |
| Pl. height | 1.069 | 0.49 | 1.300 | 0.57 | 2.081 | 1.18 | 1.742 | 0.94 |
| Ear height | 0.689 | 0.71 | 0.927 | 0.93 | 1.791 | 2.46 | 1.726 | 2.38 |
| Ear/plant | 0.011 | 0.86 | 0.013 | 1.06 | 0.022 | 2.11 | 0.022 | 2.20 |
| Moisture | 0.144 | 0.64 | 0.168 | 0.75 | 0.255 | 0.99 | 0.356 | 1.47 |
| B/L plant | 3.365 | 56.53 | 1.983 | 22.52 | 3.730 | 24.11 | 1.502 | 12.93 |
| Intensity 20 % | Syn ZP HS | Syn ZP/USA HS | Syn ZP S1 | Syn ZP/USA S1 | ||||
| Trait | ΔG | %* | ΔG | %* | ΔG | %* | ΔG | %* |
| Yield | 0.186 | 2.04 | 0.212 | 2.45 | 0.203 | 4.57 | 0.472 | 11.14 |
| Pl. height | 0.853 | 0.39 | 1.037 | 0.45 | 1.659 | 0.94 | 1.389 | 0.75 |
| Ear height | 0.550 | 0.57 | 0.739 | 0.74 | 1.429 | 1.96 | 1.377 | 1.90 |
| Ear/plant | 0.009 | 0.68 | 0.010 | 0.85 | 0.018 | 1.68 | 0.017 | 1.76 |
| Moisture | 0.115 | 0.51 | 0.134 | 0.60 | 0.203 | 0.79 | 0.284 | 1.17 |
| B/L plant | 2.684 | 45.09 | 1.581 | 17.97 | 2.975 | 19.23 | 1.198 | 10.31 |
* - expected genetic gain from selection is shown in % in relation to normal mean of studied populations
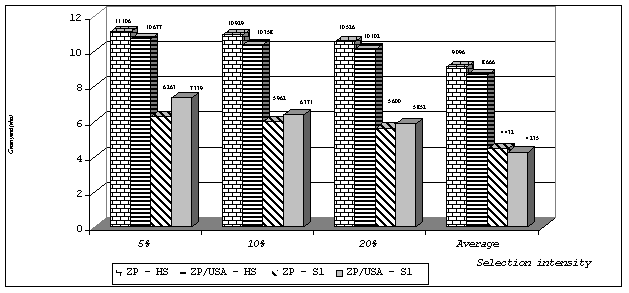
The average grain yields indicate that significant differences were obtained among observed populations. The lowest average grain yield was detected in the population ZP/USA-S1, but the average yield detected in this population at different selection intensities was always higher than the average yield recorded in the population ZP – S1 (Figure 1). Moreover, estimated genetic gain from selection was higher in the population ZP/USA-S1 than in the population ZP-S1. It is obvious that crossing of local and introduced germplasm resulted in a high concentration of favourable alleles for grain yield in a part of the material. The greatest expected gain from selection (66.44%) for stalk lodging was recorded in the population ZP-HS, while the highest gain for the grain moisture content (1.72%) was detected in the population ZP/USA-S1 at the selection intensity of 5%. Since these traits are of a great importance for the improvement of local populations it can be stated that the advantage of introduced germplasm is only fragmentary as expected progress was higher only for the grain moisture content.
Figure 1. Average grain yields (t ha-1) in observed populations at the different selection intensities
CONCLUSIONS
The introduction of introduced germplasm did not result in the improvement of grain yield, plant height, ear height or the number of ears per plant. It should be mentioned that although the average grain yield was lower in the population ZP/USA-S1, a certain number of genotypes in this population was significantly more yielding than in the population ZP-S1. On the other hand, a highly significant decrease of the grain moisture content and the percent of lodged and broken plants was obtained, which was also one of the aims of crosses to the introduced material. A three-fold higher genetic variability for grain yield was obtained by the introduction of introduced germplasm in the population ZP/USA-S1, which resulted in the increase of broad-sense heritability by 12%. Such high heritability, with exercise of a certain amount of reserve, points out to the obtained increase of frequencies of favourable alleles for grain yield in the stated population. Genetic variability and heritability of the grain moisture content was increased in the population ZP/USA-S1, which together with the fact that the average grain moisture was decreased, suggests that a significant advancement was made in such a direction. Genetic variability and heritability for lodged and broken plants were reduced, but stalk resistance to lodging was improved at the same time, which was one of the aims of the introduction of introduced germplasm. Changes in allele frequencies in the population ZP/USA-S1 initiated the increase of values of expected genetic gain from selection for grain yield. The introduction of introduced germplams resulted in the increase of genetic gain from selection for the number of ears per plant, while a positive response to selection was not recorded for other traits. Considering that a significant difference in grain yield was detected among S1 populations at different selection intensities, it is advisable to use the intensity of 10% in the further studies so that genetic variability of the material could be maintained with the positive response to selection. Based on the obtained results, the population ZP/USA can be used as a good initial material for further selection.
Albrecht B., Dudley J.W., 1987. Evaluation of four maize populations containing different proportions of exotic germplasm. Crop. Sci., 27: 480–486.
Baker R., 1984. Some of the open pollinated varieties that contributed most to modern hybrid corn. Ann. Illinois Corn Breeders School Proc., 20: 1–20.
Bletsos E.A., Goulas C.K., 1999. Mass selection for improvement of grain yield and protein in a maize population. Crop Sci., 39:1302–1305.
Bridges W.C. Jr., Gardner C.O., 1987. Foundation populations for adapted by exotic crosses. Crop. Sci., 27: 501–506.
Cochram W.G., Cox G.M., 1957. Experimental designs. John Wiley & Suns, Inc. Canada, pp 350.
Eberhart S.A., 1980. Factors effecting efficiencies of breeding methods. African Soils, 15: 669–680.
Edmeades G.O., Bolaños J., Chapman S.C., Lafitte H.R., Bänziger M., 1999. Selection improves drought tolerance in tropical maize populations: I. Gains in biomass, grain yield, and harvest index. Crop Sci., 39:1306–1315.
Efron Y., Everett H.L., 1969. Evaluation of exotic germplasm for corn hybrids in northern United States. Crop. Sci., 9: 44–47.
Geadelmann J.L., 1984. Using exotic germplasm to improve northern corn. Ann. Corn Sorghum Res. Conf. Proc., 39: 98–110.
Goodman M.M., 1984. Evaluation of exotic. Ann. Illinois Corn Breeders School Proc., 20: 85–101.
Griffing B., Lindstrom E.W., 1954. A study of the combining abilities of corn inbreds having varying propotrions of Corn Belt and non-Corn belt germplasm. Agron. J., 46: 545–552.
Hallauer A.R., 1978. Potential of exotic germplasm for maize improvement. In: Maize breeding and genetics, Walden, D.B. ed., John Wiley & Sons Inc., New York: 229–247.
Hallauer A.R., 1989. Fifty years of recurrent selection in corn. Illinois Corn Breeders School, 25:39–63.
Helms T.C., Hallauer A.R., Smith O.S., 1989b. Genetic drift and selection evaluated from recurrent selection program in maize. Crop. Sci., 29: 602–607.
Horner E.S., Lutrick M.C., Chapman W.H., Martin F.G., 1976. Effect of recurrent selection for combining ability with a single-cross tester in maize. Crop. Sci., 16: 5–8.
Lonnquist J.H., 1974. Considerations and experiences with recombinations of exotic and Corn Belt maize germplasm. Proc. Ann. Corn Sorghum Res. Conf., 29: 102–117.
Michelini L.A., Hallauer A.R., 1993. Evaluation of exotic and adapted maize (Zea mays L.) germplasm crosses. Maydica, 38: 275–282.
Nelson H.G., 1972. The use of exotic germplasm in practical corn breeding programs. Ann. Corn Sorghum Ind. Res. Conf. Proc., 27: 115–118.
Smith J.S.C., Smith O.S., Wright S., Wali S.J., Walton M., 1992. Diversity of U.S. hybrid maize germplasm as revealed by restriction fragment length polymorphism. Crop. Sci., 32: 598–604.
Sprague G.F., Eberhart S.A., 1976. Corn breeding. p.305–362. in: Corn and corn improvement, Sprague, G.F. ed., American Society of Agronomy. Madison, WI.
Trifunović V., 1983. Wide crosses in maize. Genetika, 15: 203–218.
Return to the MNL Volume 79 Index
Return to the index of Maize Newsletters
Return to the Maize Genome Database Page
{kind=link}