PIRACICABA, SP, BRAZIL
University of São Paulo, ESALQ
Maize chromosome polymorphism: A comparison between pachytene chromosomes and mitotic chromosomes using C-banding and FISH of 5S rDNA and Cent-4 probes in tropical hybrids
--Santos-Serejo, JA, Aguiar-Perecin, MLR, Bertão, MR
Pachytene chromosomes have been considered the most appropriate stage for maize cytogenetics, due to the presence of easily detectable components, such as heterochromatic knobs, centric heterochromatin and enlarged chromomeres. Accurate identification of mitotic chromosome markers is highly desirable for studies involving somatic tissues and for identification of polymorphism among maize varieties.
In this note, we report some aspects of the investigation of the karyotypes of two tropical maize hybrids by conventional pachytene analysis, C-banding of mitotic metaphases and fluorescence in situ hybridization (FISH) of a 5S rDNA probe, as a marker for chromosome 2 (Mascia et al., Gene 15:7–20, 1981), and of a chromosome 4 centromere-specific repeat, designated Cent-4 (Page et al., Genetics 159:291–302, 2001). The hybrids studied (441123 × 4443 and 441311 × 442612) were obtained in our laboratory by crossing sister inbred lines, derived from a flint maize variety (Jac Duro, Sementes Agroceres, Brazil). Their knob composition (see Table 1) is quite similar, except for chromosome 9, which is homozygous for K9S in the hybrid 441131 × 442612 and heterozygous in 441123 × 4443. In the present note, we aimed to emphasize the importance of the use of in situ hybridization of specific chromosome markers to resolve a problem of identification of chromosomes 2 and 4 in these materials.
A method previously described for root pretreatment and fixation (Bertão and Aguiar-Perecin, Cariologia 55:115–119, 2002) was employed to obtain metaphase spreads used in the C-banding (carried out as described in this reference) and FISH procedures. Preparations of microsporocytes for analysis of pachytene chromosomes were made according to current routine (Dempsey, The Maize Handbook, New York: Springer-Verlag, pp. 432–441, 1994).
Arm lengths of pachytene and C-banded mitotic metaphase chromosomes were determined from photographic prints and the chromosomes were identified according to their relative length and arm ratio, as described in the literature (Neuffer et al., Mutants of Maize, New York: Cold Spring Harbor Press, 1997; Aguiar-Perecin and Vosa, Heredity 54:37–42, 1985). Relative lengths of the mitotic chromosomes and C-bands were expressed as percent of the length of chromosome 10.
The 5S rDNA probe was a fragment amplified by PCR using the primers 5′GTGCGATCATACCAGC(AG)(CT)TAATGAACCGG3′ and 5′GAGGTGCAACACGAGGACTTCCCAGGAGG3′ (see Gottlob-McHugh et al., Genome 33:486–494, 1990). The Cent-4 probe (cloned in Bluescript and kindly provided by Dr J. A. Birchler, University of Missouri, USA) was amplified by PCR using M13 forward and M13 reverse primers. The Cent-4 sequence was labeled with biotin-14-dATP using the BioNick labeling system (Life Technologies), and the 5S rDNA probe with digoxigenin-11-dUTP, by random primed DNA labeling (Roche). In situ hybridization was performed as described (Schwarzacher and Heslop-Harrison, Practical in situ hybridization, Oxford: Bios, 2000), with slight modifications. Fluorescein-conjugated sheep anti-digoxigenin (Roche) was used to detect the rDNA sites. Three antibodies were necessary for a consistent detection of Cent-4 sequences: mouse anti-biotin, TRITC-conjugated rabbit anti-mouse and TRITC-conjugated swine anti-rabbit (DAKO). Chromosomes were counterstained with DAPI.
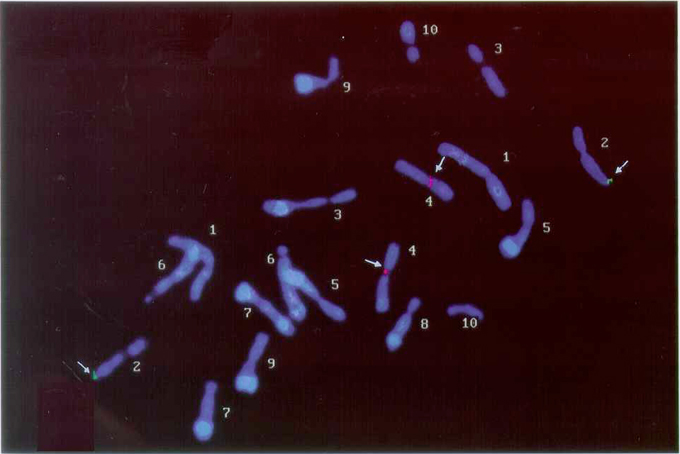
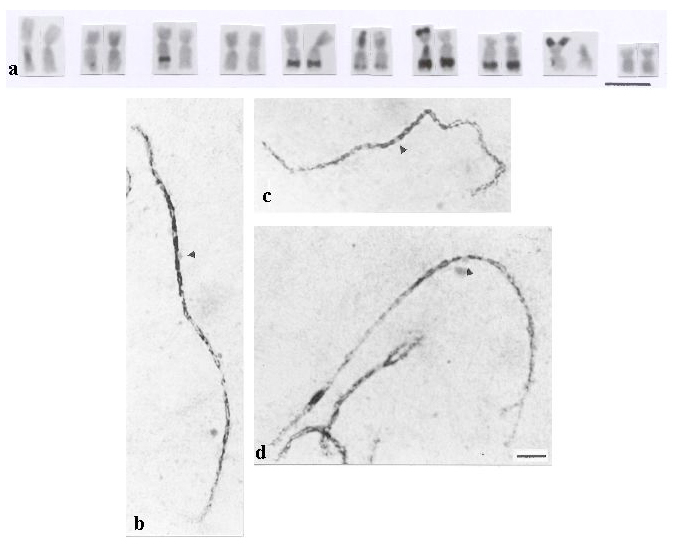
Figure 1 shows that in situ hybridization using fluorescein-labeled 5S rDNA revealed that the probe was localized on the long arm of a pair of chromosomes with submedian centromeres (arm ratio = 1.43 at mitotic metaphase and 1.71 at pachytene). The FISH signal of the TRITC-labeled cent-4 sequence was detected on the centromere of a pair of chromosomes with arm ratio values of 1.25 at mitotic metaphase and 1.30 at pachytene. Interstitial signals were not observed, as reported for other genotypes investigated by Page et al. (2001). Due to these arm ratio values and relative lengths, as well, these chromosomes had been identified as 4 and 5, respectively, in a previous paper (Bertão and Aguiar-Perecin, 2002). We reconsidered this interpretation, considering that 5S rDNA has been mapped to chromosome 2 (Mascia et al., 1981) and that the Cent-4 sequence was identified on the centromere of chromosome 4 (Page et al., 2001). Based on this conclusion, we organized Table 1, in which the knob composition, relative lengths and arm ratios of mitotic chromosomes, and arm ratios of pachytene chromosomes are shown. Figure 2a illustrates the C-banded karyotype of hybrid 441123 × 4443. Most of the values in Table 1 are coherent with the ones described in the literature, except for chromosomes 2 and 4. The arm ratio value of chromosome 2 at pachytene (1.71) seems rather high as compared with the range of 1.14–1.42 reported in the literature (Neuffer et al., 1997). Also, the range reported for arm ratio of chromosome 4 (1.63 to 2.0) is higher than our finding (1.30). The main point is that generally, the centromere is located in a position more submedian in chromosome 4 than in chromosome 2, and this is not the case in the materials investigated and also in other related lines (not shown). Figures 2b and c show these chromosomes at pachytene. Chomosome 5, which possesses a knob on the long arm (Figure 2d) is also shown, because it had been interpreted as number 2 in our previous paper. In this case, its arm ratio at pachytene (1.13) is consistent with the pattern described in the literature. As shown in Figure 2a and Table 1, in mitotic metaphases this chromosome is larger than chromosomes 2, 3 and 4 and its arm ratio is 1.35, due to the presence of the C-band in the long arm. In addition, the length and arm ratio of the other chromosomes, estimated on C-banded metaphases of hybrid 441123 × 4443, are coherent with values previously described in a comparative study between knobless chromosomes of Ceremonial race and the karyotype of a highly knobbed stock of Zapalote Chico (Aguiar-Perecin and Vosa, 1985). Also, relative lengths of C-bands are quite correspondent to knob sizes visualized at pachytene and expressed arbitrarily as large, medium and small, as seen in Table 1.
The present study emphasizes the importance of the use of molecular markers for cytogenetic analysis of maize chromosome polymorphism, that must be due not only to the presence of detectable knobs. We could speculate that variable content of different classes of repetitive could modify the gross morphology of the chromosomes as discussed here for chromosome 2 and 4, considering, for instance, that retrotransposon insertions have increased the size of the maize genome during its evolution (see San Miguel et al., Nature Genetics 20:43-45, 1998).
Table 1. Biometrical analysis of mitotic C-banded metaphase and pachytene chromosomes of the hybrid 441123 × 4443.
| Parameters | Chromosomes # | ||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | ||||
| KL+ | KL_ | KL+ | KL2+ /L3+ | KS+ / KL+ | KS-/KL+ | KL1 +/L2 + | KS+ | KS - | |||||
| Metaphase RL | 181.11 | 150.13 | 155.35 | 151.31 | 145.18 | 157.49 | 126.81 | 160.47 | 147.03 | 138.31 | 138.56 | 110.25 | 100 |
| AR | 1.18 | 1.43 | 1.82 | 1.72 | 1.25 | 1.35 | 1.75 | 1.64 | 2.38 | 2.49 | 0.88 | 1.63 | 1.74 |
| C-band RL | 15.91 | 20.36 | 12.03 | 19.89 / 27.09 | /27.09 | 21.03 | 27.29 | ||||||
| Pachytene | |||||||||||||
| AR | 1.28 | 1.71 | 2.59 | 1.30 | 1.13 | 6.73 | 2.17 | 3.82 | 1.47 | 2.68 | |||
| Knob * | M | M | S / S | M / L | M / S | L | |||||||
RL = Relative lengths of chromosomes and C-bands expressed as percent of the length of chromosome 10.
AR = Arm ratio (long arm/short arm).
* Knob size = L, large; M, medium; S, small.
#: Numbers refer to chromosomes; knob location: (+), presence of knob; (-), absence of knob; knobs at 6L2, 6L3, and 8L1, 8L2 observed at pachytene, appear as a single band on mitotic chromosomes.
{kind=link}
{kind=link}