JOHNSTON, IOWA
Pioneer Hi-Bred International
Use of anthocyanin pigmentation as a diagnostic tool to visualize drought stress in maize
--Habben, J
Visualization of drought stress in maize typically involves observing symptoms such as reduced plant height, as well as leaf rolling and leaf senescence. While these are useful traits to assess the differential response of germplasm to drought, they do not offer a tissue or cellular level understanding of the spatial and temporal response of plants to water deficits. In an effort to enhance our understanding of this facet of drought, we created transgenic maize that expresses a construct composed of a drought responsive promoter (rab17) fused to a chimeric transcription factor (CRC) whose product induces anthocyanin biosynthesis.
In maize, it is well known that rab17 responds to increased levels of ABA (Busk, PK et al., Plant J. 11:1285–1295, 1997) and that ABA accumulates in drought stressed plants (Zeevaart, JAD and Creelman, RA, Annu. Rev. Plant Physiol. Plant Mol. Biol. 39:439–473, 1988). It is also well known that anthocyanin synthesis requires regulatory genes from both the c1 and r families (reviewed in Mol, J et al., Trends Plant Sci. 3:212–217, 1998). At Pioneer, a chimeric C1 and R-paralog fusion gene (CRC) has been created and its product demonstrated to induce synthesis of anthocyanins in maize (Bruce, W et al., Plant Cell 12:65–-80, 2000). Thus, our intent was to combine these two well-characterized genetic elements to produce transgenic maize that would synthesize anthocyanins in tissues which perceive water deficits.
Ninety events containing the rab17:CRC construct were generated in a genetic background that does not normally produce anthocyanins. As an initial test of construct efficacy, intact auricles of several T0 events were incubated in either water (-ABA) or 20 μM ABA (+ABA). Within 24 hours, it was readily apparent that anthocyanin synthesis was induced in auricles exposed to ABA but not in control auricles (Fig. 1A): thus demonstrating the inducibility of the construct by this phytohormone. Approximately one week prior to anthesis, a subset of events was exposed to a water deficit (WD) while a second set of (clonal) events remained well watered (WW). Within 48 hours, anthocyanin pigmentation was observed in drought stressed leaves (Fig. 1B), stalks (Fig. 1C), and tassels (image not shown). In mature T1 kernels, pigmentation was primarily observed in the embryo and aleurone (Fig. 1D). Although embryos contained a high concentration of anthocyanin pigment, this concentration was not so great that it caused embryo lethality in subsequent generations.
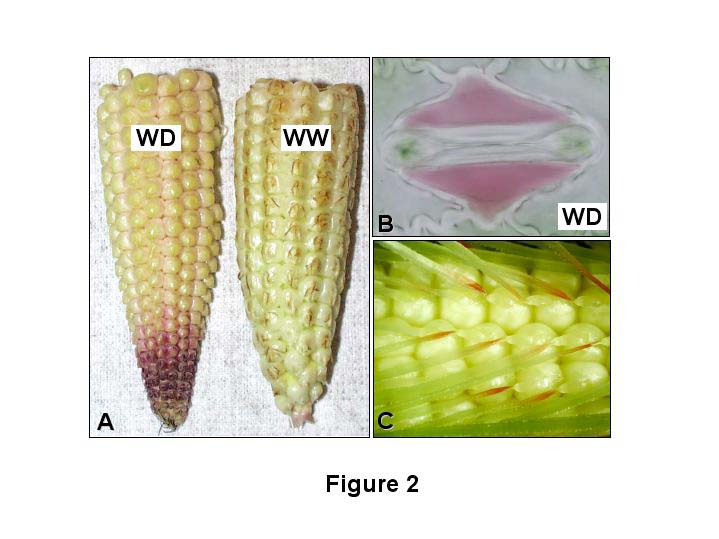
We further substantiated the utility of these transgenic events as a drought visualization tool by looking for anthocyanin deposition in organs that have previously been demonstrated to have elevated ABA levels under water deficit conditions. First, it has been shown that in developing ears exposed to drought stress there is a significant increase in ABA levels in apical kernels of drought stressed plants relative to their well watered controls (Wang, Z et al., Ann. Bot. 90:623–630, 2002). Fig. 2A shows that kernels from ear tips of rab17:CRC plants exposed to a drought stress have an increase in pigmentation relative to their well watered counterparts. Second, it is well established that ABA mediates stomatal closure in response to drought stress in plants (reviewed in Schroeder, J et al., Annu. Rev. Plant Physiol. Plant Mol. Biol. 52:627–658, 2001). We performed epidermal peels on transgenic leaves exposed to drought and well watered conditions and determined that in contrast to stomata exposed to full irrigation (image not shown), those exposed to water deficits accumulate anthocyanins (albeit in the subsidiary cells; Fig. 2B).
We also discovered anthocyanin pigmentation in cell types that, to the best of our knowledge, have not previously been noted for their accumulation of ABA. For example, Fig. 2C shows a very specific deposition of pigment adjacent to the silk abscission zone in the proximal end of post-pollinated silks. This pigmentation sector occurs independently of whether plants are well watered or drought stressed. Based on these observations, we hypothesize that the formation of the abscission zone initiates the dehydration of the silk, which in turn triggers an increase in ABA levels and subsequent activation of anthocyanin biosynthesis.
For decades, maize geneticists have exploited anthocyanin pigmentation as an in planta colorimetric assay to visualize various molecular mechanisms, thereby obviating the need for complex analytical techniques. As demonstrated here, by placing anthocyanin synthesis under the control of a well-characterized drought stress promoter, we can create a diagnostic tool that should have similar applicability to visualize the molecular mechanisms of the drought response in maize.
Figure 1. Images of anthocyanin deposition in plant parts of rab17:CRC events. A. Auricles of a T0, V5 plant incubated in either water (-ABA) or 20 μM ABA (+ABA). B. Ear leaves of T0 plants exposed to well watered (WW) or water deficit (WD) conditions. C. Cross section of stalks from T0 plants exposed to well watered (WW) or water deficit (WD) conditions. D. Longitudinal section of a mature T1 kernel; Em = embryo, Al = aleurone. Figure 2. Images of anthocyanin deposition in T1 rab17:CRC events. A. Ear tips from 11 DAP plants exposed to water deficit (WD) or well watered (WW) conditions. B. Stomata from an epidermal peel of an event exposed to water deficit (WD) conditions. C. Proximal ends of post pollinated silks (3 DAP).{kind=link}
{kind=link}