The current study involved bt1-based tropical supersweet corn inbred Hi38-71, an inbred with a high tolerance of P. polysora and of corn leaf aphids. A generation mean analysis was conducted with this inbred, rating 3.2 on a 1 to 9 scale, crossed to inbred G24 (rating 6.9). The G24 inbred derives its susceptibility from corn belt dent inbred B68, and is one of the G Set RILs from the cross of Ki14 (a Thailand inbred) and B68 (as Hawaii conversion, Hi31). This G Set of RILs segregated approximately 50% tolerant: 50% resistant (Moon et al., Maydica 44:301, 1999). Many tropical inbreds show greater resistance than Hi38-71, suggesting it to be only intermediate in mature-plant resistance. Data are summarized here from a natural epibiotic in the autumn of 2002.
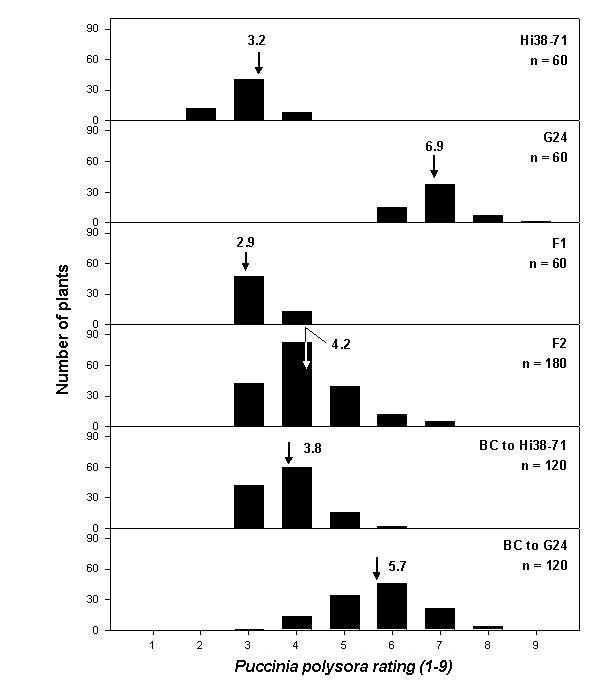
Mean ratings on the 1 to 9 scale were 2.9 for F1, vs. 3.2 for the resistant parent (Pr) and 6.9 for the susceptible (Ps) parent (Figure 1). These populations were distinguished by extremely low variances. The F2 averaged 4.2 with a broad distribution, and means of backcrosses returned toward those of the parents, 3.8 for BC to Pr and 5.7 for BC to Ps. Heterosis for vigor and yield was very great for these segregating populations, probably obscuring the genetic segregation for rust tolerance loci. For example, the F1 hybrids scored more resistant than their resistant inbred parent. There was no clear distinction of two classes in the BC to Ps.
Generation mean analysis of the 6 generations (Table 1) revealed a large mean effect but no significant additive or dominance effects. Additive x additive interaction was highly significant, and the broad-sense heritability was relatively low (31.2%). An estimate of minimum number of gene loci, using Castle and Wright formula, was 2.81.
Hi38-71 was bred from a Cuban-flint derived sh2-based sweet corn inbred, AA8sh2, that carries resistance to the corn leaf aphid, Rhopalosiphon maidis (Fitch) (Chang and Brewbaker, MNL 48:37-38, 1974). We have confirmed this aphid resistance to be recessive and monogenic, as noted by Lu and Brewbaker (MNL 73:36, 1999). AA8sh2 was converted to Rp1-d, derived from W22xB14A, previously suggested to be linked to the aphid resistance locus (Chang and Brewbaker). It appears possible that one or more of the Southern rust resistance loci in Hi38-71 are linked to the aphid and Rp1 (common rust) loci
Holland et al. (Theor. Appl. Genet. 96:232-241, 1998) reported major QTLs for mature-plant resistance to southern rust on chromosome 10S, with minor QTLs on chromosomes 3 and 4. The 10S locus accounted for 83% of the variation in resistance in their F2:3 populations of tropical x Corn Belt. The hypernecrotic southern-rust resistance locus, Rpp9 (Ullstrup, A.J., Plant Dis. Rep. 34:89-99, 1965) is closely linked on Chromosome 10 to gene Rp1 for resistance to Puccinia sorghi Schw. The common sources of Rpp9 confer little or only partial tolerance to southern rust races in the tropics. We concur with Holland et al. that a few major loci undergird general resistance to southern rust in tropical germplasm, and that Rpp alleles may also serve in this capacity.
Table 1. Estimates of additive, dominance, and interaction parameters
for the Hi38-71 x G24 family.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
am=midpoint, a=additive, d=dominance, aa=additive x additive interaction, ad=additive x dominance interaction, dd=dominance x dominance interaction
Figure
1. Frequency distribution of P. polysora rating on 6 generations.
Return to the MNL 77 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}