During the 2000 field season, we monitored the level of cell wall phenolics and particularly diferulates in inbred lines of maize developed by Agriculture and Agri-Food Canada with improved resistance to Gibberella ear rot (CO432 and CO433). Twelve days after silk emergence, plants of this inbred plus susceptible inbreds were artificially inoculated with a macroconidial suspension of F. graminearum. Control plants were inoculated with distilled water. Ears were collected at 0, 2, 4, 6, 8, 10, 15, 20, 30 days and harvesting time. Ergosterol levels were monitored by HPLC (Young, J. Agric. Food Chem., 43(11), 2904-2910, 1995) in whole grains. Pericarp and aleurone layers were separated from the rest of the grain. Cell wall bound phenylpropanoids were analyzed after NaOH digestion of crude cell wall using a HPLC and LC-MS that we developed.
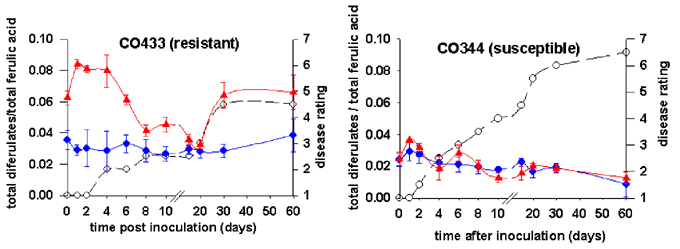
The levels of diferulates were significantly different among genotypes. More interestingly, in CO433, a resistant genotype, diferulates/ferulate ratio was significantly higher in inoculated pericarps than in the controls during the first six days of infection and after the twentieth day (Figure 1). No such differences were found in CO344, a susceptible genotype. High levels of diferulates impede the action of cell wall hydrolases (Grabber et al., J. Sc. Food Agric. 77:193-200, 1996). The high level of dimerization of the cell wall may impede cell wall degradation and cell disruption necessary for the progression of Fusarium which is a necrotroph phytopathogen. Cross-links of cell wall proteins in response to fungal elicitors have already been described in bean and soybean cell suspension (Bradley et al., Cell 70: 21-30, 1992) and is considered as a quick and early response of the plant to pathogens.
Figure
1. Diferulates/ferulate ratio in maize pericarp of infected (s)
and control (l) pericarp and aleurone
layers. Disease ratings (m) are also
shown (lower values indicate less infection).
Return to the MNL 76 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}