In the genus Zea, the chromosome number of all the species is 2n=20 except Zea perennis which is 2n=40. For quite some time, Zea perennis has been considered an autotetraploid and other species have been thought to be diploid species. But much evidence supporting x=5 as the basic chromosome number of the genus Zea, which was proposed by Anderson (Chron. Bot. 9:88-92, 1945), has been obtained (Molina and Naranjo, Theor. Appl. Genet. 73:542-550, 1987). Anderson pointed out that the genus Zea could be originated by allopolyploidy from different diploid ancestral species (2n=10), probably extinct at the present time. In these diploid species, the distinct original genomes (x=5) that are present in the actual polyploid may be differentiated. It is most important to know the basic number of the Zea genus for clarifying the hypothesis proposed by Anderson.
The highly conserved tandem repeats 5S and 45S rDNA sequences are useful chromosome markers for understanding genomic evolution and taxonomic relationships at both the molecular and the chromosomal levels. They have been successfully applied for phylogenetic analysis in many other plants (Jiang and Gill, Chromosoma 103:179-185, 1994; Fukui et al., Theor. Appl. Genet. 96:325-330, 1998; Sang and Liang, Genome 43:918-922, 2000).
In this study, we have detected the number and distribution of 5S and 45S rDNA loci in three teosinte species including Zea mays ssp. mexicana (2n=20), Zea diploperennis (2n=20) and Zea perennis (2n=40), by FISH, and provide additional information for their phylogenetic relationships.
Plant materials. The materials tested include Zea mays ssp. mexicana (2n=20), Zea diploperennis (2n=20) and Zea perennis (2n=40). The materials were kindly supplied by Professor M. G. Gu (Institute of Genetics, Chinese Academy of Sciences, Beijing)
DNA Probes and labelling. The plasmids of 5S and 45S rDNA which were cloned in the vector pUC18 (Arumuganthan et al., Mol. Gen. Genet. 22:551-558, 1994) were kindly provided by K. Arumuganthan, University of Nebraska. The tested probes were labeled with Bio-11-dUTP following the nick translation protocol using the kit (Sino-American Biotechnology Company, China. See also Gustafson and Dille, 1992). Dot Blotting was performed to detect the labelling efficiency.
Chromosome preparation. The root tips of 1-2 mm length from vigorously grown teosinte plants were treated with saturated bromonaphthalene at 25 C for 2.5 h, and fixed with methanol-acetic acid (3:1) overnight. After being fully washed with water, the root tips were treated with a mixture of 2% pectinase and 2% cellulase (SERVA) at 28 C for approximately 3 h. Finally, the treated root tips were squashed and dried in a flame (Song and Gustafson, Genome 36:658-661, 1993). The slides were kept in a -20 C freezer for FISH.
In situ hybridization and fluorescence detection (FISH). For in situ hybridization, the method published by Gustafson and Dille (Natl. Acad. Sci. USA 89:8646-8650, 1992) was adopted with some modifications. Slide preparations were pretreated with 100µg/ml RNase in 2xSSC (0.3 M sodium chloride plus 0.03 M sodium citrate) at 37 C for 1 h, rinsed briefly in 2xSSC, probe DNA was then denatured by immersing the slides in 70% deionized formamide in 2x SSC at 70 C for 3.5 min. After dehydration in an ice-cold ethanol series, 40µl of denatured probe mixture (50% (v/v) formamide, 10% (w/v) dextran sulfate, 0.5% (w/v) SDS (sodium dodecyl sulfate), 0.2% (w/v) sheared salmon sperm DNA, in 2xSSC) was added to each slide and covered with a coverslip. Hybridization was carried out at 37°Ê in a humid chamber overnight. The slides were then washed in 2XSSC, 0.1XSSC, at 42 C for 3-5 min each step, and subsequently washed in 0.1% of TritonX-100, PBS at RT, 5 min each step. Sites of hybridization were detected with streptavidin-Cy3 (Pharmacia) at 37 C for 30 min. Finally the slides were counterstained with 1 µg/ml DAPI. Chromosomes were viewed with an Olympus BX60 fluorescence microscope equipped with a Sensys 1401E CCD camera. Red and blue images were captured in black and white with different filters. The images were combined and pesudocolored in the computer using software V++.
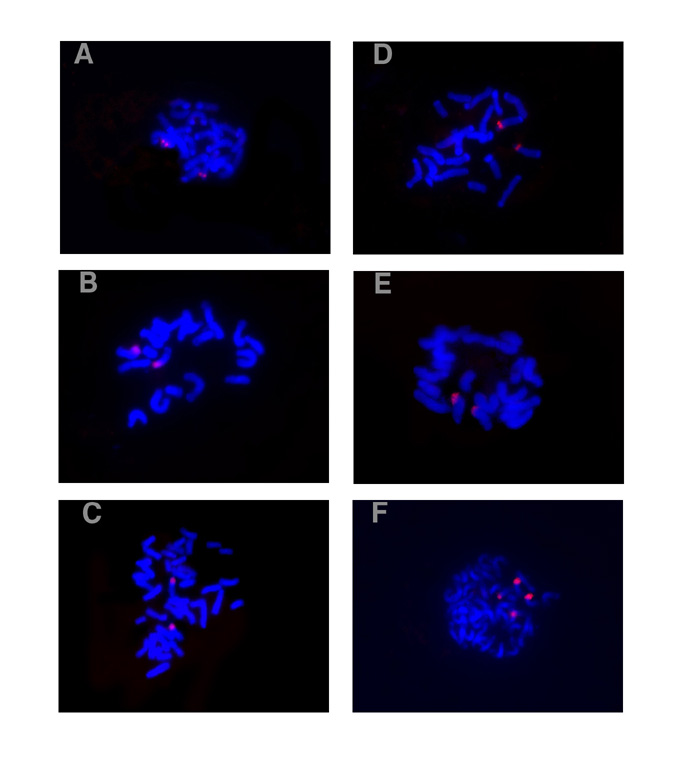
The FISH results showed that the chromosomes were blue, while the signals were red (Fig. 1). Each of the tested materials showed only one hybridization site of 5S rDNA and all of the sites were located on the subterminal regions (Fig. 1A-C). It was detected on the long arm of chromosome 2 in Zea mays ssp. mexicana (Fig. 1A), on the short arm of chromosome 5 in Zea diploperennis (Fig. 1B), and on the short arm of the detected chromosome pair, which could not be identified, in Zea perennis (Fig. 1C). The detected chromosomes for Zea mays ssp.mexicana and Zea diploperennis were recognized according to the standard karyotypes published by Zhang et al. (J. Wuhan Univ. 1:111-132, 1992) and Pasupuleti et al. (J. Hered. 73:168-170, 1982) respectively. In Zea mays ssp. mexicana and Zea diploperennis, the signal spot of 45S rDNA was detected on the subterminal region of the short arm of chromosome 6 where it covered the nucleolar organizer (NOR) and no minor 45S rDNA loci were detected (Fig. 1D-E). Whereas in Zea perennis, hybridization signals of 45S rDNA were localized on the subterminal regions of the short arms in two different chromosome pairs (Fig. 1F). The signal spots were larger on one pair of them, and smaller on the other. The signal spots for two members of each detected homologous chromosome pair resemble each other in size. It meant that the differences of signal size between the two detected chromosome pairs were not induced by technical reasons. They seem to correspond to the major and minor locus for 45S rDNA respectively.
The signals were observed on each chromatid in the detected chromosomes, which showed separated sister chromatids (Fig. 1A, D and E), while the larger signals combined by two spots were detected in those in which each two sister chromatids had not separated yet (Fig. 1B, C and F).
Our results showed that Zea mays ssp. mexicana and cultivated maize (Zea mays) (Mascia et al., Gene 15:7-20, 1981) were the same for the position of 5S rDNA, which were located on the long arms of chromosome 2, while Zea diploperennis and Zea perennis were different and their positions of 5S rDNA were located on the short arms of the unidentified chromosomes. These results support the biochemical, karyotypic, and morphological evidence for the suggested taxonomy in the genus Zea.
Based on synapsis data, Molina and Naranjo (1987) deduced that all species of the genus Zea were allotetraploid except for Zea perennis. RFLP mapping showed that many markers mapped to two or more chromosome locations in the maize genome (Davis et al., Genetics 152:1137-1172, 1999). Actually, different genes in the maize genome could have single, double, triple, and even quadruple locations according to ISH results (Li et al., Hereditas 129(2):101-106, 1998; Ren et al., Hereditas 126(3):211-217, 1997). This means that only a part of the genes were tetraploid in the maize genome. It is evident that synapsis analysis could not tell the site number for a given sequence, nor did the Southern hybridization techniques, the copy number of which could only be inferred. However, FISH technique could not only confirm the site number of the gene on chromosomes, but also map them to a special region. In this study, the FISH results of the location for 5S rDNA showed that all of the tested Zea species had only one site in their genomes. For 45S rDNA, all also showed one site except for Zea perennis, which had two sites on different chromosomes. All of the sites were located on the subterminal regions of the chromosomes. Compared with the RFLP linkage map, the FISH results were also more precise. Therefore, FISH technique should be a powerful tool for the analysis of structure in a genome.
Zea perennis showed two 45S rDNA hybridization sites correspondingly to two different chromosome pairs (Fig. 1F). This provides evidence that Zea perennis is not a simple autopolyploid; it is allotetraploid at least for 45S rDNA. The results were also supported by the results with meiotic and karyotypic analyses (Molina and Naranjo, 1987).
If the genus Zea really originated as an allopolyploid from different diploid species (2n=10), that probably are extinct at the present time, as has been postulated (Anderson, 1945), then our results clearly demonstrated that diploidization and evolutionary loss of the 5S rDNA and 45S rDNA have apparently occurred in three teosinte species. This is the case because each diploid ancestral genome must have at least one site of 5S and 45S rDNA in their genome. Such evolutionary reorganization of rDNA loci has been reported in several plant genera in the studies with allopolyploid species including Scilla (Araki, Genetica 66:3-10, 1985; Vaughan et al., Heredity 71:574-580, 1993), Milium (Bennett and Thomas, Genome 34:868-878, 1991) and Brassica (Maluszynska and Heslop-Harrison, Ann. Bot. 71:479-484, 1993). The mechanisms for the diploidization and evolutionary loss of genes remain to be studied.
We would like to thank Prof. K. Arumuganthan, University of Nebraska
for kindly providing the 5S and 45S rDNA probes. The work was supported
by the National Natural Science Foundation of China (No. 30070376) and
the Research Fund for the Doctoral Program of Higher Education (No. 207980112).
Return to the MNL 76 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}