Little is known about the regulation of type I DNA-topoisomerase (topo I) activity in plant mitochondria. We have previously reported (MNL 69:63-64, 1995; MNL 70:29-30, 1996; MNL 71:40-41, 1997; MNL 72:33, 1998) on the effect of different redox conditions (including the redox system of glutathione) on mitochondrial genome expression regarding DNA, RNA and protein synthesis in organello. Significant activation of transcription and translation in mitochondria under oxidising conditions and its profound repression under reducing conditions can indicate the existence of a special mechanism of redox regulation of genetic functions in plant mitochondria.
The aim of the present work was to verify our hypothesis that mitochondrial topo I is a redox regulatory enzyme which is regulated by the glutathione system via the reduction/oxidation of a critical disulfide bridge(s). Three-day-old etiolated maize seedlings of hybrid VIR42 MV were used for mitochondria isolation. The method of topo I purification was the same as described in our note from this issue. Topoisomerase activity was measured as previously described (Tarasenko and Konstantinov, Biopolymers and Cell 14:111-116, 1998).
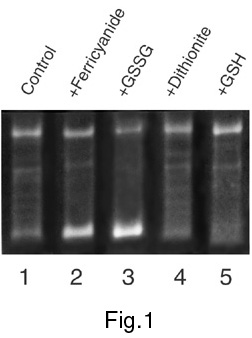
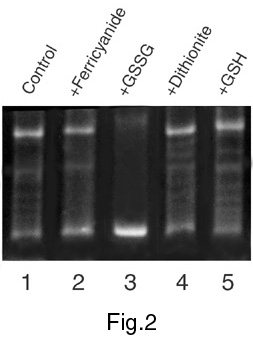
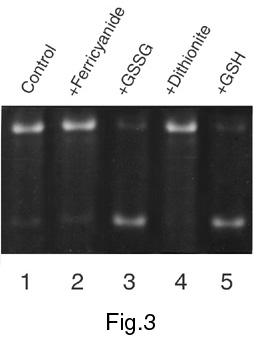
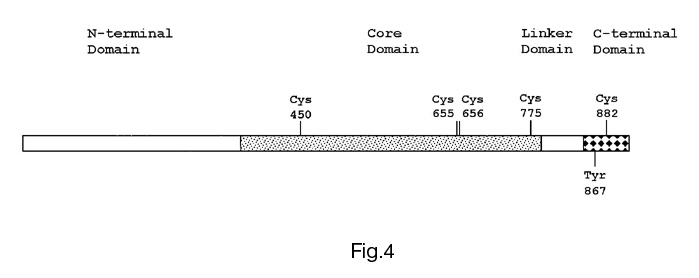
The relaxation activity of mitochondrial topo I has been studied under model changes of redox conditions created by the addition of potassium ferricyanide and oxidised glutathione (GSSG) as oxidising agents, and sodium dithionite and reduced glutathione (GSH) as reducing agents. The addition of both oxidising and reducing agents causes the changes in the activity of topo I from mitochondria (Figures 1-3) but the effects of redox agents depend strongly on the stage of enzyme purification. Figure 1 shows an activation of topo I in the presence of dithionite or GSH and its significant repression following the addition of ferricyanide or GSSG after the stage of ammonium fractionation. As regards the effects of these agents on topo I activity after the stage of chromatography on the column with DEAE-Toyopearl ("Toyosoda", Japan), the effects were observed just for oxidised and reduced forms of glutathione (Figure 2). Studying of the redox agents effects on the topo I activity after the final stage of enzyme purification (Figure 3) revealed that both GSSG and GSH caused inhibition of relaxation activity. As a whole, the results of these experiments show the dependence of mitochondrial topo I activity on redox conditions. These results also suggest the existence in mitochondria of special regulatory enzymes involved in reduction/oxidation of cysteine residues in the topo I protein molecule. Figure 4 shows the location of cysteine residues in the amino acid sequence of plant topo I potentially involved in redox regulation of topo I activity.
The data presented demonstrate that type I DNA-topoisomerase from maize mitochondria has the properties of a redox regulatory enzyme.
Financial support from the INTAS (Project Number 98-0522) is acknowledged.
Figure 1. The effect of redox conditions on topoisomerase activity after ammonium sulfate fractionation.
Figure 2. The effect of redox conditions on topoisomerase activity after chromatography on DEAE-Toyopearl.
Figure 3. The effect of redox conditions on topoisomerase activity after chromatography on dsDNA-cellulose ("Pharmacia", USA).
Figure
4. The domain structure of the plant type I DNA-topoisomerase. The
scheme was made on the basis of data of Balestrazzi et al., Gene 183:183-190,
1996. Cys - cysteine, Tyr - "active site" tyrosine. Numbers designate amino
acid positions.
Return to the MNL 73 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}