The disease induced by infection of Helminthosporium turcicum is one of the serious maize diseases and is distributed in many countries including China. It damages maize production seriously. It was reported that there are four Helminthosporium turcicum resistance loci, namely ht1, ht2, ht3 and htn1 (Zaitlin et al., 1992). The ht1 was first identified by Hooker (1963). Recently, the genetic working map accompanying the RFLP map in maize was published (Coe, 1995). In this map, many genes including ht1 were placed at the coordinate locations defined in the RFLP map. The gene ht1 is 168 cM from the zero point and linked closely to umc22 with 4.3 cM on one side and umc122 with 1.2 cM on the other side. However, the genetic map could only show the relative locations and distances of genes or markers, but physical maps show real positions and distances of genes or markers on the chromosomes. Up to now, very little work has been reported concerning physical mapping of genes of agronomic interest in maize (Shen et al., 1987; Hong et al., 1993; Song et al., 1997; Li et al., 1997). The physical location of the gene ht1 of maize has not been reported yet, as we know.
In situ hybridization (ISH) technique is a powerful tool to physically locate specific DNA sequences or genes directly on chromosomes. The sensitivity of in situ hybridization (ISH) techniques for humans is now down to 0.25kb (Richard et al., 1994). The ISH of small single or low copy DNA sequences in plants was more difficult than in humans. Recent developments and numerous refinements of methods including chromosome preparations and detection procedures have made it possible to detect signals of small single or low copy DNA sequences in plants (Gustafson et al., 1990; Gustafson et al., 1992; Song and Gustafson, 1995; Song et al., 1997; Li et al., 1997; Ren et al., 1997). In these studies, the signals were detected only by DAB. Recently, fluorescence in situ hybridization (FISH) has developed rapidly in plants. However, the probes used in these reports were either total genomic DNA (Mukai et al., 1993; King et al., 1994), repetitive DNAs (Griffor et al., 1991; Pederson et al., 1995; Linares et al., 1996) or large DNA fragments (Ambros et al., 1986) or BAC clones (Jiang et al., 1995). Only Dong and Quick (1994) mapped a 2.6 kb single/low copy DNA sequence on metaphase chromosomes of wheat and rye by FISH. It was demonstrated that FISH had many advantages over ISH using DAB detection method, for example, high sensitivity and higher contrast between signals and chromosomes (Jiang et al., 1994; Heiskanen et al., 1996). The FISH of DNA segments around 1 kb in size has never been reported so far in plants. The study to detect and map smaller single or low copy DNA sequences and genes by FISH is very important for physical mapping of plant genomes.
RFLP markers are very good landmarks for mapping genes. Even though the gene ht1 has not been cloned, its physical location could be deduced by the physical position of the RFLP markers linked closely with it. In this study, we have determined the chromosome location of ht1 by FISH of the two RFLP markers umc22 and umc122 linked tightly to and flanking it.
Plant and RFLP marker materials: Maize (Zea mays L.) inbred line Huang Zao 4 (Li, 1997), which derived from a native cultivar in China, was used as the tested plant material, the seeds were provided by Professor Song Jianchen, Shandong Agricultural University, Shandong Province, P.R. China. Both tested RFLP markers umc22 and umc122 are close to the ht1 gene dictating reaction to Helminthosporium turcicum on the long arm in linkage group 2 of the classic genetic maps (Coe, 1995). They are 780 and 450 bp DNA fragment size cloned in pUC19 and kindly offered by the RFLP laboratory and probe bank of the U. S. Department of Agriculture, Agriculture Research Service, and Department of Agronomy, University of Missouri, Columbia, USA.
Chromosome preparation: Chromosome preparation methods were developed using the protoplast technique as described by Song et al. (1995) with some modifications. Root tips were collected from germinating seeds and treated in a-bromonaphthalene for 2 hr at RT (room temperature), then washed in deionized water for 15 min. The root tips were fixed in freshly prepared methanol:acetic acid (3:1) fixative overnight at 4 C, subsequently washed in deionized water for 15 min. Then the root tips were digested in 1% cellulase (Shanghai Institute of Biochemistry, Chinese Academy of Sciences) and 1% pectinase (SERVA) at 28 C for 2.5 hr. After enzymatic treatment, cells were subjected to a hypotonic treatment in water, then 2-3 root tips were squashed with forceps in 2-3 drops of fixative on a cold slide and flame dried. The prepared slides were stored at �20 C or immediately used for FISH.
Biotin labeling DNA and in situ hybridization: The probe was biotin-labeled with the procedure of the kit supplied by Sino-American Biotechnology Company China. After labeling reaction at 15 C for 2.5 h, labeling was stopped by adding 5 ul 0.2M EDTA (pH 8.0). Then, labeled probe was separated through a Sepharose column and evaluated by means of dot blots.
In situ hybridization was performed using the procedure described by Gustafson and Dille (1992).
Detection: Fluorescent signals were detected based on the procedure published by Griffor et al. (1991), Gustafson and Dille (1992) and Dong and Quick et al. (1994) with some modifications. The detection reagents came from a kit offered by Beijing Medical Academy. The procedure includes the following steps: 1) after hybridization, the coverslip was removed by dipping the slides in a 2 x SSC solution and the slides were washed in 20% formamide (in 2 x SSC) at 42 C for 10 min, 2 x SSC at 42 C for 10 min, 0.1% Triton at room temperature for 4 min and PBS at room temperature for 5 min, then the slides were drained but not dried; 2) 20 µl FITC-avidin D was added to each slide, the slides were covered with a coverslip, incubated in a humid chamber at 37 C for 30 min and the slides were washed 3 times, each for 5 min in PBS at room temperature; 3) 20 µl of biotinylated goat anti-avidin D antibody were added to the slides for 30 min at 37 C in a humid chamber, then slides were washed as above; 4) repeat step 2; 5) after washing in 2 x SSC at room temperature for 5 min and subsequently in PBS at room temperature for 5 min, 10 µl (1 µg/ml) propidium iodide in an anti-fade (10 µg/ml) solution were added to the slides, the slides were covered with a coverslip, and observed and photographed under a Zeiss fluorescence microscope.
An average of the hybridization site measurements was taken by calculating the distance from the centromere to the detection site and using that as a percentage of the arm on which the site was located. The arm ratio of the chromosome showing a detection site was also measured in order to determine on which chromosome the site was located.
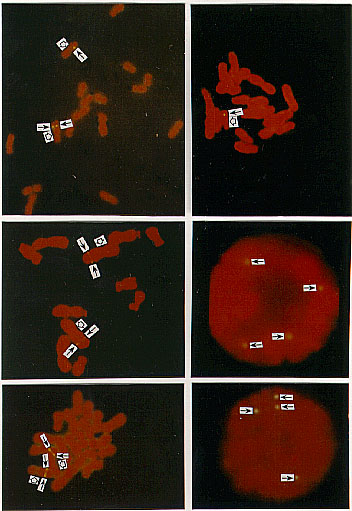
Counterstained by FITC and propidium iodide (PI), the chromosomes appeared red and the signals showed yellow. The hybridization signals of both umc22 and umc122 were showed on 2L (the long arm of the chromosome 2) and their percentage distances from the hybridization site to the centromere were 61.78 and 63.01 respectively (Figs. 1a, c and e). The signals of umc22 and umc122 were also detected on 7L and the percentage distances were 47.54 and 48.85 respectively (Figs. 1b, e). In some cases the detected cells showed only one signal spot on one chromatid of one member of the detected chromosomes (Fig. 1b); but in another case, the detected cells usually showed signals on both homologous chromosomes or two sister chromatids of one member of the homologous chromosomes (Figs. 1a, c). In some detected cells, the signals from both sister chromatids were merged into a large one, just like a band (Fig. 1e). Interphase nuclei were also scored in our FISH analysis. In 100 observed interphase cells, about 60 cells showed signals. Signals were shown on 1, 2, 3, and 4 sites of different interphase nuclei respectively. Figures 1d and f showed the nuclei hybridized by umc22 and umc122 respectively, and each had 4 hybridization sites. The karyotypes showing the genetic and physical map of umc22 and umc122 were presented in Figure 2 so as to compare them.
Figure 1a-f. The hybridized sites of the probes umc22 and umc122 on the chromosomes in maize. In all figures, the black arrows denote the hybridization signals, the white arrows denote the centromeres. a) The hybridization signal of umc22 is in 2L. b) The hybridization signal of umc22 is in 7L. c) The hybridization signal of umc122 is in 2L. d) The interphase cell has four hybridization signals of umc22. E) The hybridization signal of umc122 is in 2L and 7L. f) The interphase cell has four hybridization signals of umc122.
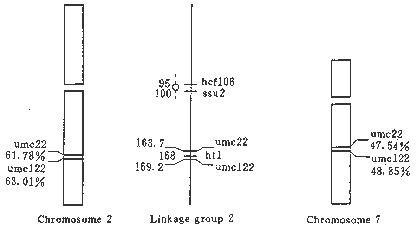
Figure 2. The idiograms of the locations of umc22 and umc122 markers on the chromosomes and the comparison of their sites between the chromosomes and the genetic linkage groups.
The probes umc22 and umc122 both showed two different hybridization sites on 2L and 7L. It means that they are duplicated sequences. It has been demonstrated that many of the RFLP markers were duplicated at least once somewhere else in the maize genome. That maize is an allotetraploid was proved by different maize geneticists (Poggio et al., 1990; Tito et al., 1991olina et al. 1992). Some scientists demonstrated that the duplicated sequences of maize RFLP markers probably were relevant to properties of the allotetraploid (Weber, 1990; Dowty and Helentjaris, 1992; Song et al., 1997). It can be thought there were some homologous sequences or regions in the different original species from which maize originated even though they belonged to different genomes. Helentjaris et al. (1988) noted that the maize RFLP marker duplications were not necessarily scattered randomly over the entire chromosome, but more often appeared to cluster. Dowty and Helentjaris (1992) reported that there was also a higher order pattern to the duplication saturated regions of three different "sets" of chromosomes: 10-2-7, 5-1-9 and 3-8-6. For example, chromosome 2 shares duplications with both chromosomes 7 and 10, chromosomes 7 and 10 do not share any duplicated markers. Helentjaris (1995) further reported that the regions, 2L with 7 etc. shared extensive similarity. In this study, our results were just consistent with the above reports. The markers umc22 and umc122 not only share the same chromosome arms, 2L and 7L, but also have almost the same percentage distances between them (1.2 on 2L and 1.3 on 7L), even the order of these two markers and centromeres was the same on both 2L and 7L. It demonstrated that different ancestor species from which maize originated had strong conservativeness in the sequences, organization and order of markers in genomes or regions of the genomes.
The gene ht1 (Helminthosporium turcicum resistance1) is linked closely to umc22 with 4.3 cM on one side and umc122 with 1.2 cM on the other side. Because genetic and physical maps show the same marker order, the gene ht1 should be physically located between the hybridization sites of umc22 and umc122, i.e., percentage distances 61.78 and 63.01 on 2L. Our results demonstrated that both umc22 and umc122 had their homologous sequences on 7L besides 2L and the order of these two markers and centromeres was the same on both 2L and 7L. Therefore, the gene ht1 probably also had its homologous sequence between two hybridization sites of umc22 and umc122 on 7L. Actually, the signal spot showing the hybridization site of the marker umc122 probably also represents the physical location of ht1, because each signal spot could occupy about 1-3 cM.
Our results demonstrated that FISH has a number of advantages over DAB (diaminobenzidine tetrahydrochloride) detected ISH. First, the yellow shown by the hybridization signals has high contrast with the red displayed by the chromosomes, while in the DAB detected ISH it is more difficult to distinguish the color not only between the signal and chromosomes but also between the signal spots and those contaminated by Giemsa staining. Second, for small single or low copy DNA hybridization, FISH can get higher detection rate and find more detected cells showing signals on two members or two sister chromatids of one member in the homologous chromosomes. In this study, their rates were about 15%, while the detection rates were only about 10%. Very few or even no cells in which the signals were detected simultaneously on two homologues or two sister chromatids could be found in DAB ISH (Gustafson et al., 1990; Gustafson et al., 1992; Song and Gustafson, 1995; Song et al., 1997; Li et al., 1997; Ren et al., 1997). Third, the markers at multiloci can be located simultaneously by multicolor FISH and it is very useful for confirming the order of closely linked genes or markers on the chromosomes. However, FISH, as a powerful tool of molecular cytogenetics, has not been reported so far for mapping of single or low copy markers around 1 kb in size in plants, not as in animals. We think the main obstacle is chromosome preparation. The protoplast chromosome preparation techniques developed by Gustafson and Dille (1992) and modified by Song and Gustafson (1995) can get rid of the cell debris hiding the chromosomes and offer more target chromosomes for combining of the probes. This will be beneficial to improving hybridization efficiency and compensating the shortage of low detection rate for small single or low copy DNA hybridization. Therefore, one of the effective approaches to develop FISH techniques in plants is combining FISH with protoplast chromosome preparation.
In ISH studies of rice RFLP markers it was indicated that considerable variation could, and did, exist between genetic and physical maps (Gustafson and Dille, 1992; Song and Gustafson, 1995). In the genus Triticum it has been demonstrated that the physical distances between genes are often quite different from the genetic distances (Lukaszewski and Curtis, 1993). Heslop-Harrison (1991) thought there was often little correlation between the separation distances of markers on the genetic and physical map. However, in this study, the tested markers umc22 and umc122 located at the middle part of the long arm in linkage group 2 of the genetic map (Coe, 1995), according to our results are also physically located at the middle part of 2L. The genetic distance between these two markers was 5.5 cM, the percentage distance was 1.2 and the distance in both genetic and physical maps was close to each other. It demonstrates that the maize physical map is basically consistent with the genetic map for umc22 and umc122. At present, the physical location of most genes or markers is still unknown. We suggest that different species and genes should not be the same for the relationship between the genetic and physical map. Although there are variations between these two types of map, the correlation between them will be displayed for more and more genes or markers as their physical mapping is developed.
Acknowledgements: The tested RFLP markers offered by Dr. Coe�s
laboratory, U. S. Department of Agriculture, Agricultural Research Service,
Plant Genetics Research Unit and Department of Agronomy, University of
Missouri. This research was supported by the Natural Science Foundation
in China and the Doctorate Vesting Point Foundation of the Education Committee
of the People�s Republic of China.
Return to the MNL 73 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}