Eukaryotic genomes contain one to several tandem arrays of rDNA which encode for ribosomal subunits usually located in the nucleolar organizing region (NOR). The region of the nucleus where transcript processing takes place has the characteristic appearance of a dark, usually spherical object, the nucleolus.
The number of nucleoli in a cell can vary from one to as many as the number of NORs encoded in the genome. The maize gnome has one NOR on the short arm of chromosome 6, which can result in nuclei with one or two nucleoli. Morgan reported that of 519 premeiotic interphase cells, all had one nucleolus, and that of 508 cells in mitotic interphase 32 had two nucleoli (Morgan, Cytologia 36:669-673, 1971). It has also been shown that the number of nucleoli can be dependent on the cell type. Nucleoli are useful cytogenetic markers that allow the number of copies of chromosome 6 to be determined during interphase and prophase. Any study of nuclear organization must consider the number and size of nucleoli because nucleoli occupy space and, as a result of the possibility that multiple NORs could participate in the formation of one nucleolus, the organization of the nucleus could be altered. As part of such a study, we wanted to determine the effect that an additional NOR would have on nuclear organization.
The nucleoli of maize can be detected in cell spreads with a silver staining method (Howell and Black, Experientia 36:1014-1015, 1980). In order to screen for trisomic 6 plants the silver staining method was applied to intact tissue (roots and the coleoptile). In addition to identifying trisomic 6 plants, the tissue preparations revealed that the order of fused/unfused nucleoli in files of cells formed a pattern. We recognized the utility of the modified protocol for any situation where large numbers of plants or cells must be screened for changes in the number or size of nucleoli. The following modified protocol can be used to quickly stain and visualize nucleoli in root and coleoptile tissue. Tapetal and callus cell spreads were also examined using the original protocol.
Tissue was fixed in 3:1 (ethanol:acetic acid) until needed for staining. The fixative was rinsed out of roots by immersion of the tissue in water for thirty minutes. Roots were digested in citrate buffer containing 4% pectinase (v/v) for 1 to 2 hours, at 37 C, then rinsed in water for ten minutes. Next, cells were spread in 3:1 fix on a microscope slide. This produces files of cells. The slide was stained with one part developer and two parts silver nitrate at 70 C (Howell and Black, Experientia 36:1014-1015, 1980) until nucleoli could be visualized under a light microscope. The coverslip was removed by rinsing in water and the slide was dried overnight, then mounted in premount. Fixed coleoptiles were rinsed for five minutes in water, placed on a microscope slide and cut in half. Tissue was stained as above without digestion, sealed under a cover slip and visualized under a light microscope. This allowed files of cells to be scored as above. Excess stain can be removed from microscope slides by adding more developer at one side of the cover slip and drawing it across with a piece of filter paper. Microscope slides were sealed with nail polish and examined. Fresh callus (not fixed) was digested in citrate buffer containing 4% pectinase (v/v) and 5% cellulase (w/v), spread out on a microscope slide and stained the same way as roots. Tapetal cells were removed from fixative, placed in water for 30 minutes and then digested in 4% (v/v) pectinase and 5% cellulase (w/v) for 1 hour. After digestion cells were spread on a microscope slide, dried over night, silver stained and scored.
Files of stained coleoptile or root cells were scored sequentially for the number of nucleoli per cell. A length of 20 to 50 cells was counted per file and several slides were analyzed for each tissue type. Spread callus and tapetal cells were counted at random for the number of nucleoli per cell, as the cells could not be retained in files.
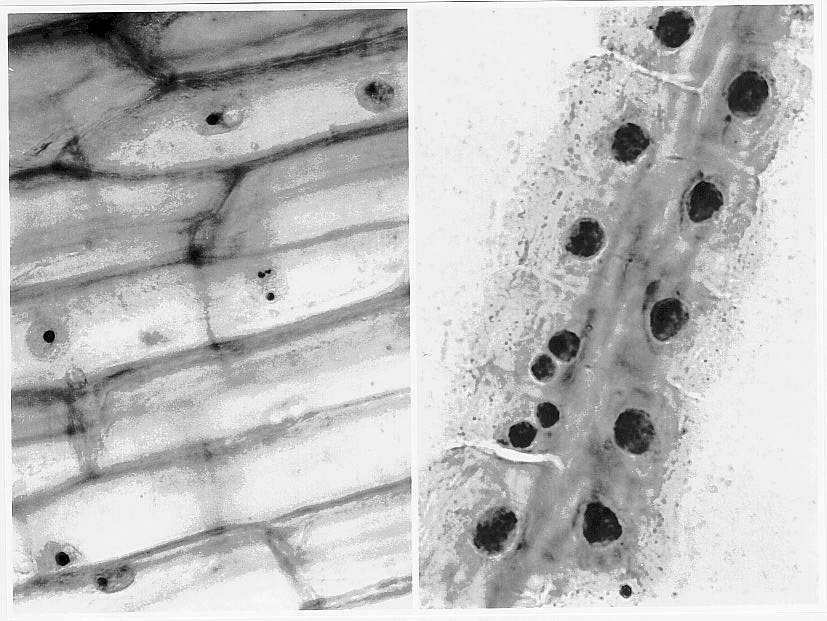
The frequency of one or two nucleoli was determined and a pattern was identified; cells containing two nucleoli are preferentially near each other in root, tapetal and coleoptile cells (refer to Table 1). Figure 1a and b show silver stained coleoptile and root cell preparations. In most cells the number of nucleoli was correlated with the size of the nucleoli: in trisomic cells there could be three small nucleoli, one small plus a medium sized nucleolus, or one large nucleolus. In the diploid cell-types the nucleus had either one medium sized nucleolus or two small nucleoli.
Genetic background and tissue-type appear to be factors affecting the frequency of fused nucleoli (see Table 1).
This phenomenon of clustered cells with the same number of nucleoli can be seen in all the tissue-types and strains of maize that we studied. It is possible that daughter cells retain positional memory from their parent cell leading often to similar/same nuclear organization of the short arm of chromosome 6. Similarly, cell to cell communication could be occurring that results in neighboring cells having similar numbers of nucleoli. Further study is underway to identify the frequency and mechanism(s) of nucleolar fusion.
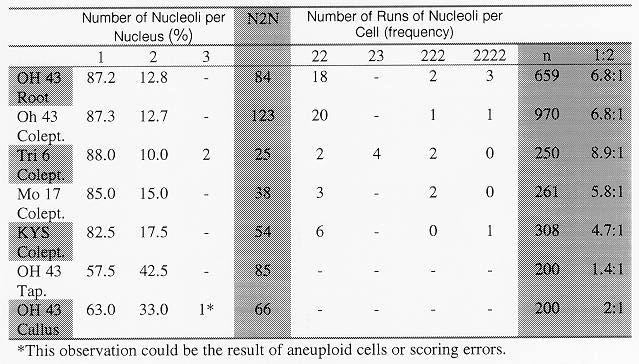
Table 1. Percentages of cells with one nucleolus (1), two nucleoli (2), or three nucleoli (3); two cells next to each other in a file of cells with two nucleoli each (22), two cells next to each other, one with three nucleoli and one with two nucleoli (23), three cells in a row all with two nucleoli (222) and four cells in a row all with two nucleoli (2222); the number of nuclei with 2 nucleoli (N2N); the total number of cells scored (n); and a ratio of cells with two nucleoli to cells with one (1:2). Abbreviations used in column one are Oh 43 (Ohio 43), Colept (coleoptile), Tri 6 (Trisomic 6), Mo 17 (Missouri 17), KYS (Kansas Yellow Saline), Tap (metaphase tapetal cells).
Figure 1. (a) Trisomic 6 coleoptile tissue, (b) Ohio 43 root file preparation (silver stained).
Return to the MNL 72 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}