The major quantitative trait loci for plant stature, development
and yield are general manifestations of developmental gene clusters
--Khavkin, E, Coe, EH
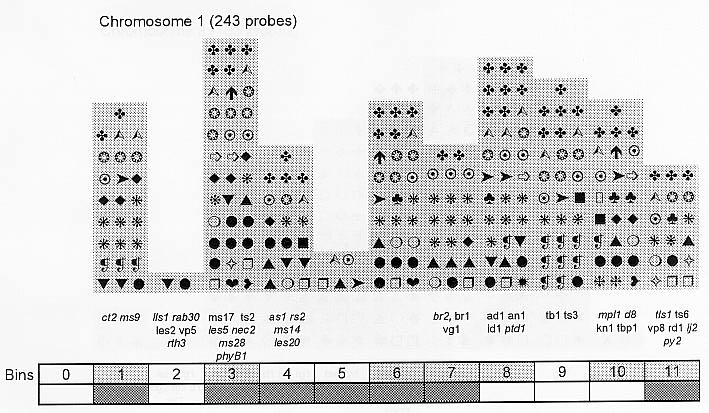
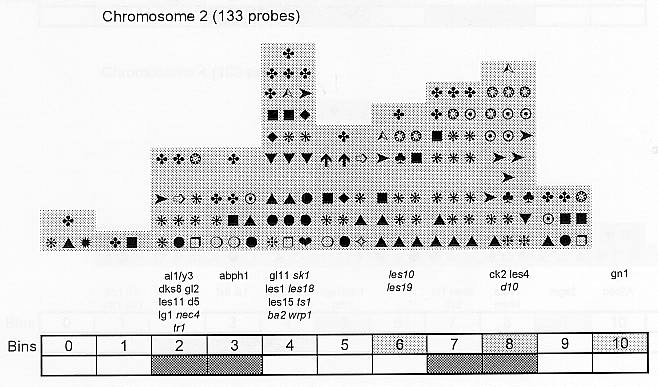
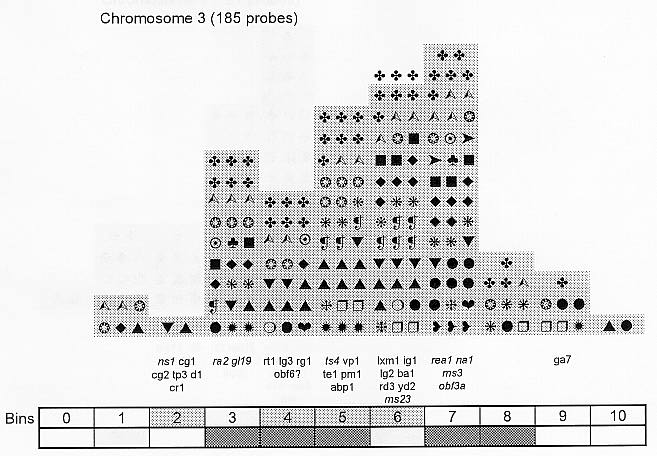
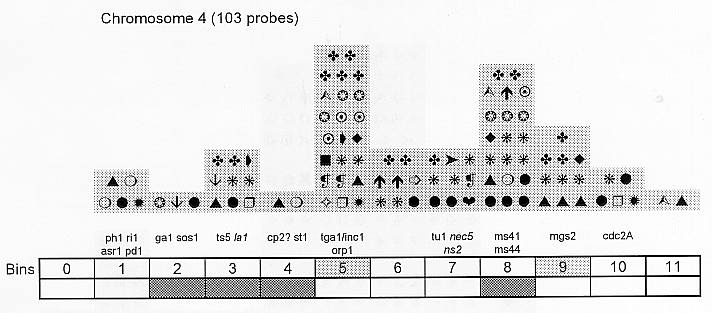
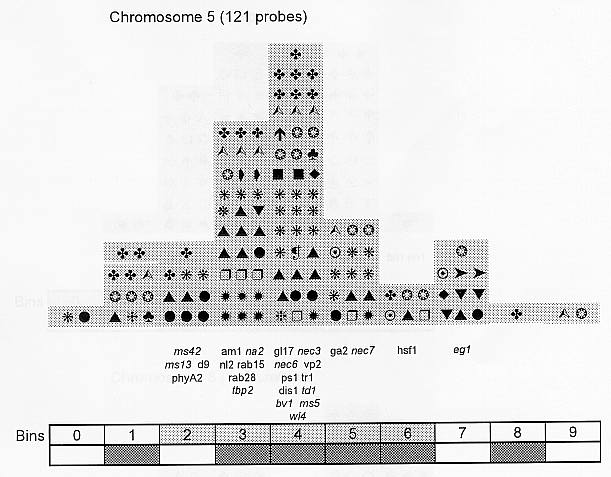
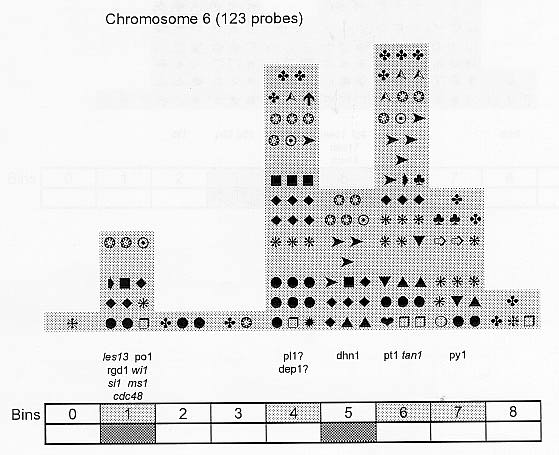
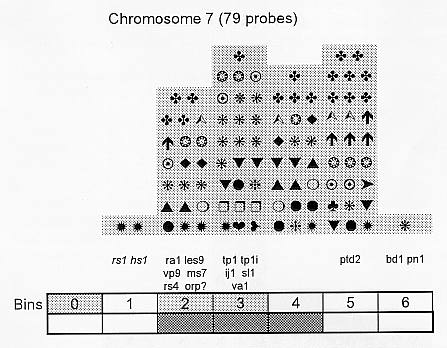
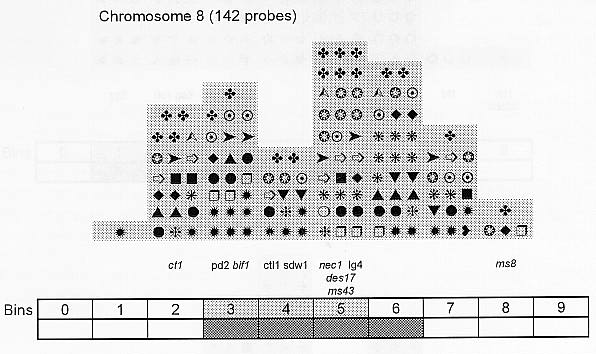
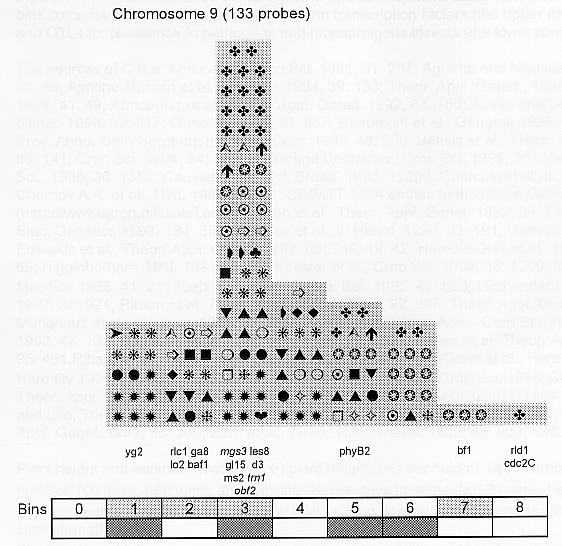
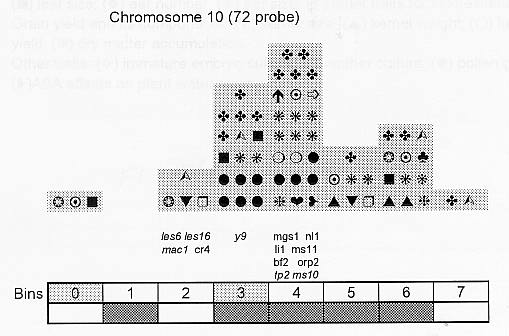
Previously (MNL 68:61, 1994; MNL 69:106, 1995; MNL 70: 42, 1996; Theor. Appl. Genet. 95: 343, 1997) we discussed functional associations of corn developmental genes. We revised the database of quantitative trait loci (QTLs) for maize plant architecture, growth and development rates in vivo and in vitro, yield and its components, and ABA accumulation and effects extracted from the current publications to conjecture QTL positions within the bins on the 1995 UMC molecular map. In this way QTLs reported by different authors were collated. Within a particular mapping population, highly diverse physiological traits were mapped by one and the same RFLP probe, whereas one and the same developmental trait was usually mapped to several positions on different chromosomes. The locations for one and the same developmental trait varied considerably in different mapping populations. When QTL distribution was followed along maize chromosomes, well-defined peaks were found. However, this pattern of QTL distribution was not specific as to putative physiological functions ascribed to particular loci. When QTLs were compared with the qualitative loci mapped by mutations and with cDNA-derived probes, such collation did not support the indirect candidate-gene approach. Taking into consideration the mapping accuracy, QTL associations coincide with the previously defined clusters of the qualitative developmental genes (Fig. 1), particularly the homeotic genes expressing transcriptional regulators. We believe that these clusters are the functional networks of genes expressed in concert to control plant development. Apparently it is these networks of the developmental genes that are phenotypically manifested as the major QTLs for plant structure, earliness and grain yield.
Many qualitative and quantitative loci affecting corn resistance to abiotic and biotic stress: drought, phytophagous insects, viral and fungal diseases (Beavis et al., Crop Sci. 34:882, 1994; Dingerdissen et al., Mol. Breed. 2:143, 1996; Lebreton et al., J. Exp. Bot. 46:853, 1995; McMullen and Simcox, Mol. Plant-Microbe Interact. 8:811, 1995; Saghai Maroof et al., Theor. Appl. Genet. 93:539, 1996; Schon et al., Heredity 70:648, 1993; Zehr et al., MNL 68:110, 1994) map within the clusters of developmental genes. This evidence presumes that the cooperative expression of developmental clusters might mediate non-specific plant responses to unfavorable environment.
Figure 1. Distribution of QTLs of plant growth and development along corn chromosomes. Shaded bins comprise the homeotic genes expressing transcription factors (the upper row) and the genes and QTLs for resistance to pathogens and phytophagous insects (the lower row). Chromosome 1,Chromosome 2, Chromosome 3, Chromosome 4, Chromosome 5, Chromosome 6, Chromosome 7, Chromosome 8, Chromosome 9, Chromosome 10.
The sources of QTLs: Abler et al., Crop Sci. 31:267, 1991; Agrama and Moussa, Euphytica 91:89, 1996; Ajmone-Marsan et al., Maydica 39:133, 1994, Theor. Appl. Genet. 90:415, 1995, Maydica 41:49, 1996; Armstrong et al., Theor. Appl. Genet. 84:755, 1992; Austin and Lee, Theor. Appl. Genet. 92:817, 1996, Genome 39:957, 1996; Beaumont et al., Genome 38:968, 1995; Beavis, Proc. Annu. Corn Sorghum Ind. Res. Conf. 49:250, 1994; Beavis et al., Theor. Appl. Genet. 83:141, 1991, Crop Sci. 34:882, 1994; Berke and Rocheford, Crop Sci. 35:1542, 1995; Bohn et al., Crop Sci., 36:1352, 1996; Causse et al., Mol. Breed. 1:259, 1995; Charcosset et al., MNL 68:44, 1994; Chernov A.A. et al., MNL 68:49, 1994; CIMMYT 1994 entries to the Maize Genome Database (http://mnl.maizegdb.org); Cowen et al., Theor. Appl. Genet. 84:720, 1992; Doebley and Stec, Genetics 134:559, 1993; Doebley et al., J. Hered. 85:191, 1994, Genetics 141:333, 1995; Edwards et al., Theor. Appl. Genet. 83:765, 1992; Heredia-Diaz et al., Maydica 41:65, 1996; Higginbotham, MNL 65:65, 1991; Koester et al., Crop Sci. 33:1209, 1993; Kozumplik et al., Maydica 41:211, 1996; Lebreton et al., J. Exp. Bot. 46:853, 1995; Lubberstedt et al. Crop Sci. 37:921, 1997; Murigneux et al., Genome 37:970, 1994; Phillips et al., Proc. Annu. Corn Sorghum Ind. Res. Conf. 47:135, 1992; Ragot M. et al., Crop Sci. 35:1306, 1995; Rebai et al. Theor. Appl. Genet. 95:451, 1997; Ribaut et al., Theor. Appl. Genet. 92:905, 1996, Theor. Appl. Genet. 94:887, 1997; Sari-Gorla et al., Heredity 69:423, 1992, Heredity 72:332, 1994; Schon et al., Heredity 70:648, 1993, Crop Sci. 34:378, 1994; Stuber et al., Theor. Appl. Genet. 132:823, 1992; Veldboom et al., Theor. Appl. Genet. 88:7, 1994; Veldboom and Lee, Theor. Appl. Genet. 89:451, 1994, Crop Sci. 36:1310 & 1320, 1996; Wan Y., Theor. Appl. Genet. 85:360, 1992; Zehr et al., Theor. Appl. Genet. 83:903, 1992, MNL 68:110, 1994.
Return to the MNL 72 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}