Development of PCR-based markers to facilitate large-scale screening
in molecular maize breeding
--Hu, X; Ribaut, JM; González de León, D
RFLPs are the most commonly used and informative molecular markers in cereal genome mapping and have been used to locate genes governing both qualitative and quantitative traits. RFLP analysis, however, requires relatively large quantities of DNA, and once the DNA has been transferred onto membranes by Southern blotting, the analysis is cost-effective only if these membranes are reprobed multiple times. In marker assisted selection (MAS) experiments, however, a subset of individuals may be selected after genotyping a single locus of interest and all individuals that do not carry the desired allele at that locus can be eliminated; at each subsequent selection step at other loci, the population is reduced further. This can only be achieved effectively by using PCR-based marker systems as described in Ribaut et al. (Plant Mol Biol Rep, submitted). Here we describe the set of PCR-based markers used in an ongoing MAS experiment for the improvement of drought stress tolerance in a CIMMYT tropical maize line.
Conversion of RFLP markers to PCR-based sequence tagged sites (STSs) offers several advantages over conventional RFLP analysis. These include the relative ease and rapidity of PCR reactions, the potential for rapidly screening very large populations (> 1000 individuals), the convenience of sharing primer sequences, and the relatively small amount of fairly crude genomic DNA needed for amplification (Erpelding et al., Genome 39: 802-810, 1996). More recently, PCR-based microsatellite or simple sequence repeat (SSR) markers have been developed in several crops (Roder et al., Mol Gen Genet 246: 327-333, 1995 Saghai Maroof; et al., PNAS 91: 5466-5470, 1994 Wu and Tanksley, Mol Gen Genet 241: 225-235, 1993), as an alternative method to detect DNA polymorphisms. In maize, many SSR markers have been identified and mapped, and more than 200 SSR primer sets are available for public use (Taramino and Tingey, Genome 39: 277-287, 1996 MaizeDB, 1996). The major limitation of STS- and SSR-PCR analysis is that it requires extensive sequence information in order to design appropriate primers.
A number of loci at QTL peaks detected in mapping experiments involving a donor and a recipient line were investigated for the development of PCR-based markers for the transfer of the corresponding genomic segments from the donor to the recipient line.
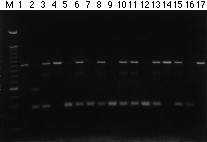
A set of 9 pairs of primers was synthesized based on existing sequence information for eight RFLP probes (Table 1), and were evaluated using genomic DNA of the tropical maize lines Ac7643S5 (donor) and CML 247 (recipient). Among nine STS-PCR primer sets, only the primer set for umc67 sts amplified polymorphic products between the two lines at an annealing temperature of 56 C without subsequent restriction digestion (Fig. 1). One fragment (~1000 bp) was amplified from Ac7643S5 and two (~600 bp and ~400 bp) from CML247. Tests of individual F2 plants showed that the 1000 bp fragment from Ac643S5 and the 400 bp fragment from CML247 could be consistently scored as a codominant STS locus marker (Fig. 1). Because the ends of the PCR products are defined by the primer sequences, the amplified length polymorphism produced by the primer set for umc67 sts must represent insertions/deletions in the DNA between the primer sequences. The rest of the primer sets amplified monomorphic products, single or multiple copy, between the two lines. The single or two copy products of primer sets for umc32, umc49, bnl7.08, and bnl7.49 were digested with ten 4- or 5-base-recognizing restriction enzymes (AvaII, HaeIII, HhaI, HinfI, RsaI, AluI, MspI, TaqI, MboI, and MseI). Polymorphisms were observed between the two maize lines when the PCR products of the primer sets for umc32 and bnl7.08 were digested with HaeIII and HhaI, respectively. However, none of the digestions of PCR products of the primer sets for umc49, bnl7.08, and bnl7.49 revealed any polymorphisms between the two lines. The low level of polymorphism detected by STS markers is in sharp contrast to the high levels detected by the corresponding clones when used as RFLP probes and demonstrates at least one important limitation of STS markers. Indeed, PCR amplification detects polymorphisms only within the region between the primer sequences, while hybridization-based RFLP analysis detects polymorphisms beyond the hybridization site of the probe. The STS-PCR primer sets which revealed polymorphisms between the two maize lines before or after enzyme digestion were used to screen 72 F2 individuals from the cross of the two maize lines. As expected, most markers revealed loci that co-segregated with the corresponding RFLP loci. The umc67 patterns in particular, which did not require digestion, were easy to score and reliable, and were used in MAS of the genomic segment marked by locus umc67. The map positions of umc32 sts and bnl7.08 sts could not be determined because the polymorphic bands produced by these two primer sets were faint and difficult to score; better primer sets might need to be designed in such cases.
Microsatellite or SSR markers provide another powerful tool to detect polymorphisms at the DNA level through simple PCR amplification. Six published SSR primer sets, detecting loci at positions of interest, were used to amplify the genomic DNA from the same two maize lines; five of these amplified polymorphisms between the two lines. All five polymorphic SSRs were mapped to positions expected from published data (Taramino and Tingey, 1996, Genome 39: 277-287; MaizeDB, 1996). Although it may be safer to verify the location of the corresponding loci in any given cross, SSR loci seem to be conserved among different maize lines. The increasing availability of SSR primer sequences for maize loci and their seemingly higher polymorphism as compared to STS markers, should make them ideal tools for large-scale PCR-based MAS experiments. However, due to the small size of their amplification products, SSR marker polymorphisms may need to be resolved by polyacrylamide gel electrophoresis, which is more time consuming and technically demanding than simple agarose separation, or by electrophoresis using high resolution agaroses (such as Metaphor), which are significantly more expensive than regular ones.
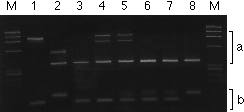
For cost-effective MAS from a large population, rapid and reliable techniques that require small amounts of DNA, less time and low-cost inputs involved in PCR reactions and gel electrophoresis, are preferred. We developed a technique for simultaneous amplification of more than one STS and/or SSR polymorphism in a single PCR reaction the products of which can be visualized in the same gel or in different gels by loading part of the amplification products from the same PCR reaction. Two factors are critical for success. First, different primer sets should be able to share common PCR amplification conditions for effective co-amplification of their respective products; finding the appropriate annealing temperature is therefore crucial. Second, the chosen markers should yield products that differ in size if effective scoring of the corresponding loci is to be achieved. After testing a range of annealing temperatures, it was found that for most double combinations of SSR and/or STS primer pairs, 56?C would give adequate or better results. Figure 2 shows the PCR products amplified with primer sets for umc67 sts and MAG1A01 in a single PCR reaction, and separated in 3% agarose gel (2% MetaPhor and 1% SeaKem). Since there was no size overlap in the respective products of each primer set, the polymorphisms corresponding to each of the two loci of interest could be easily scored in the same agarose gel. Because STS products are usually large and thus do not overlap with SSR products in size, STSs are very useful in combination with SSRs in multiplex PCR reactions in the same reaction tube. The PCR-products from the same reaction can be separated in the same or different gels depending on the size difference of the polymorphic fragments. Two people can comfortably screen two loci (whenever these can be visualized in the same electrophoretic gel) in a population of more than 2000 individuals in less than a week using this simple methodology. The cost-effectiveness, time-saving and large scale applications of this simple PCR-based methodology are obvious when compared with hybridization-based methods such as RFLP analysis. Eventually, allele-specific amplicons, giving simple presence/absence polymorphisms, and not requiring gel-based detection methods should prove to be the long-awaited method of choice for MAS from large populations. In any case, once the population size has been trimmed down to those individuals bearing the genomic segments of interest from the donor parent, the screening of other genomic regions for selection against the donor genome can then be most effectively achieved using RFLP analysis.
Table 1. Sequences of STS primers based on clone sequence data obtained
from MaizeDB.
| Clone | Locus chrom. location | Primer
name |
Sequence (5'-3') |
| umc67 | 1 | umc67F | GCTTGCTTGCCGCTAATA |
| umc67R | GACCCTGATGTTCTCCTG | ||
| umc49 | 2 | umc49F | TATAATCTCTTTGTTCTTTTT |
| umc49R | CTTCCTTTGACCACCACCTTG | ||
| umc124 | 8 | umc124F | AGATCGCCGCCGCCTACACG |
| umc124R | GCCTCTGCCACAATGACACG | ||
| umc32 | 8 | umc32F | CGGCCAGGGGTTTCTTAGG |
| umc32R | TGGCTCTTTGACATCTTCC | ||
| bnl7.08 | 8 | bnl7.08F | AAACTGGCATCATAAACATC |
| bnl7.08R | ATACAAAAGCATCAACAAGC | ||
| umc64 | 10 | umc64F1 | ATGTTGTCGCCTGCTGTGC |
| umc64R1 | AAACTTATTGTCTTCTCCA | ||
| umc64F2 | ACTTCCGTGTCTGTTTCCAA | ||
| umc64R2 | CTGATGTTCTCCTGCTGCTA | ||
| umc130 | 10 | umc130F | AGGGGGAAGAAGGTCATCAT |
| umc130R | GGCAGAGGAGGGGGAAGGAG | ||
| bnl7.49 | 10 | bnl7.49F | CCCGATGAAACAATGAAACT |
| bnl7.49R | CGGAGAGGATGGTGATGAGC |
Fig. 1. PCR amplification of genomic DNA of maize inbred lines Ac7643S5 (lane 1), CML247 (lane 2), and 15 F2 plants (lanes 3-17) from the cross of these two lines with a primer set for umc67 sts. M, size marker of 100 bp ladder. Amplification products were fractionated on 1.5% SeaKem agarose gel in 1X TBE buffer.
Fig. 2. PCR amplification of genomic DNA of two maize inbred lines Ac7643S5 and CML247, and 6 F2 plants from the cross of these two lines with two primer sets (for umc67 sts and MAG1A01) in the same PCR reaction. M, HaeIII- size marker. The amplification products were fractionated in 3% MetaPhor agarose gel in 1X TBE buffer.
Return to the MNL 71 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}