An SEM study of inflorescence development in Tripsacum dactyloides
--Orr, A; Kaparthi, R; Dewald, C; Sundberg, M
Although most current workers favor the hypothesis that maize arose from a teosinte-like ancestor, the debate gained renewed vigor with a recast of a Mangelsdorf hypothesis that Tripsacum played a role in the evolution of maize (Eubanks, Econ. Bot. 49:172-182, 1995). Fertile F1 Tripsacum-teosinte hybrid plants, that were derived when Eubanks crossed Zea diploperennis and Tripsacum dactyloides, produced hybrid inflorescences (ears) with fused, non-disarticulated cupules. According to Eubanks these first generation hybrid ears exhibit an intermediate morphological stage toward the evolution of the maize ear. If hybridization between Tripsacum and Zea played a role in the evolution of the maize inflorescence, it is likely that Tripsacum shares a similar pattern of inflorescence organogenesis with Zea (maize and the teosintes).
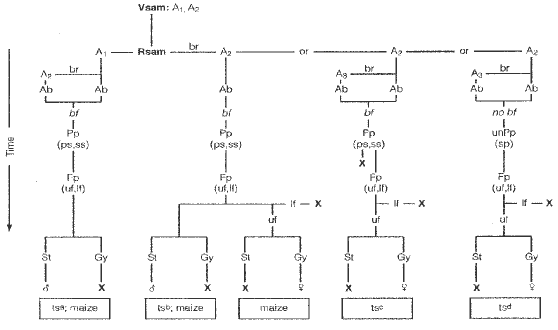
In previous studies by two of us (Orr and Sundberg) we described the development of inflorescences (ears and tassels) in the teosintes (see review Flowering Newsletter 18:48-53, 1994), and primitive maize (Am. J. Bot. 82:64-74, 1995; 83:1255-1265). The organogenic observations from these SEM studies were used to examine the prevailing ideas on the origin of the maize ear. These observations (Fig. 1) disclosed that: (1) ears and tassels in the genus Zea are derived on a common developmental background with bifurcation of spikelet pair primordia giving rise to paired sessile and pedicellate spikelet primordia; (2) the solitary spikelets of teosinte ears result from arrested growth and subsequent abortion of the pedicellate primordia; (3) the switch from 2-ranked to 4-ranked (distichy to polystichy) could be due to a change in the developmental timing program that controls the bifurcation of auxillary buds; and (4) a proposed switch from staminate to pistillate condition may have involved an intermediate bisexual (mixed) infloresence stage.
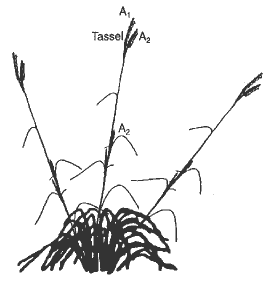
In light of Eubank's hypothesis indicating a possible role of Tripsacum in the origin of the maize ear, it became imperative that the early organogenesis of T. dactyloides bisexual inflorescences be investigated to test the developmental relationship among Zea and Tripsacum. Gamagrass plants (WW 1379) were grown at the USDA, ARS Southern Plains Research Station, Woodward, OK, and inflorescences were dissected from primary axes (A1) and lateral (A2) axes (Fig. 2). Material was prepared for scanning electron microscopy as described in previous work (Orr and Sundberg Am. J. Bot. 81:598-608, 1994).
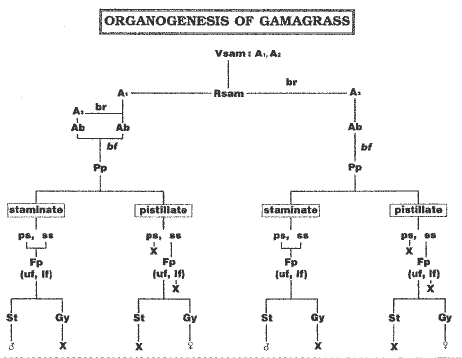
Organogenesis along the infloresence proceeded acropetally and showed a dorsiventral morphology. Distichously produced bract primordia subtended axillary buds along the abaxial surface. The development of axillary bud (ab), spikelet pair primordia (Pp), and lower floret (lf) and upper floret (uf) primordia were similar to that described for maize (Cheng et al., Am. J. Bot. 70:450-462, 1983) and the teosintes (Fig. 1). Inflorescences developed two ranks (four rows) of staminate spikelet pair primordia (Pp) with two florets each on the distal portion of the rachis, and two ranks (two rows) of single female spikelets with one floret on the proximal portion of the rachis (Fig. 3). Gynoecial tissue aborted in both staminate florets. The single spikelet condition arose by arrest and abortion of the pedicellate spikelet (ps), and abortion of the lower floret in the sessile spikelet (Fig. 3). This ontogenetic derivation of the single, sessile spikelet is similar to the pattern observed in the lateral (A2) teosinte inflorescence (Z. perennis, Z. mexicana and Z. parviglumis) (Fig. 1). The sexually mixed condition is similar to the teosinte, Zea diploperennis (Sundberg and Orr, Am. J. Bot. 73:1699-1712, 1986).
SEM analysis indicates that members of the genera Zea and Tripsacum share a common ontogenetic pattern of inflorescence ontogeny. We are continuing to characterize the development of Tripsacum inflorescences by investigating with SEM the organogenesis of a T. dactyloides gynomonoecious mutant, GSF-1.
Figure Abbreviations: Ab, axillary bud; bf, bifurcation; br, branch; Fp, floret primordium; Gy, gynoecium; If, lower floret; Pp, spikelet pair primordia; ps, pedicellate spikelet; Rsam, reproductive shoot apical meristem; ss, sessile spikelet; st, stamen; uf, upper floret; unPp, unpaired spikelet primordium; Vsam, vegetative shoot apical meristem; letters indicate primordia initiated; X, subsequently aborted.
Figure 1. The genus Zea is represented by maize, and four teosinte taxa comprising two annuals and two perennials. Generalizations in the illustration may not reflect environmentally induced variability. A1, main axis; A2, first order lateral branch off A1; A3, second order lateral A2.maize, Z. mays ssp. mays; ts, teosinte: (a, ssp. diploperennis, perennis, mexicana and parviglumis; b, ssp. diploperennis, mexicana and parviglumis; c, ssp. perennis, mexicana and parviglumis; d, ssp. diploperennis. Adapted from Orr and Sundberg, Am. J. Bot. 81:598-608, 1994.
Figure 2. Morphology of gamagrass. The tassel and ear positions on the A1 and A2 axes are shown. A1 represents main culm terminated by a sexual mixed tassel. A2 represents a sexually mixed lateral branch of the tassel or a sexually mixed inflorescence lateral to an A1 culm. Illustration modified from Jackson, 1990.
Figure
3. Comparative summary of early organogenic inflorescence development
on A1 and A2 axis of Tripsacum dactyloides. A1, main axis;
A2, secondary axis of A1.
Return to the MNL 71 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}