Epigenetic programming of paramutant R allele expression with
light and temperature conditions applied at a specific stage of seedling
development
--Mikula, BC and Kappen, T
R. A. Brink reported in 1956 that paramutation at the r locus contradicted a basic assumption of Mendelian genetics: alleles emerge from heterozygotes unchanged. The regularity of the phenomenon of paramutation has made it possible to challenge another assumption of Mendelian genetics: that environmental conditions have no heritable effect on gene expression. Under paramutagenic conditions all R alleles from a heterozygote with R-st are heritably changed. A weakly paramutagenic R-st allele reduces R pigment expression to an intermediate level of variegation. Since all R alleles are changed under paramutagenic conditions, the regularity of paramutation presented threshold conditions which made it possible to show that environmental conditions can cause heritable changes in the expression of a particular allele. Compared with the inflexibility of standard Mendelian genes, there was, with paramutation at the r locus, a high probability of being able to assay the environmental influence on paramutant R allele expression in a single generation in an inbred line and thus avoid segregating modifier arguments. We reported in MNL 67, 68 and Genetics 140:1379-87 conditions and times during early development when different levels of paramutation could be programmed. In MNL 66 we reported testcrosses from early pollen samples of plants, which as seedlings received specific light and temperature conditions, showed higher levels of paramutation (more weakly variegated) than those pollen samples made seven days later from the same plant.
The differences in paramutation between early and late pollinations
suggested that a gradient of paramutation could be found if pollen was
sampled from a single plant over the duration of anthesis, usually seven
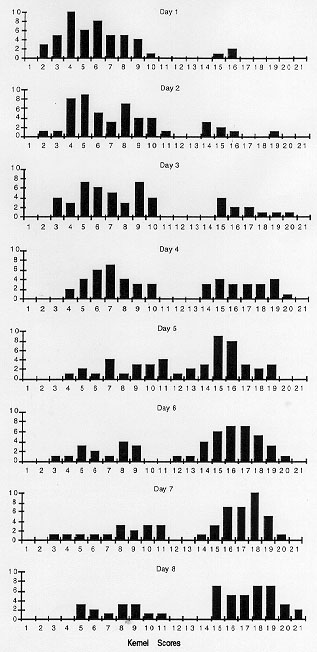
to eight days. Figure 1 shows frequency histograms for pigment scores of
kernels from testcrosses of a single plant made each of the eight days
that pollen was shed. 50 kernels from each of the eight testcross ears
were scored for the level of R pigment variegation by matching each
kernel against a set of 20 standard kernels. The frequencies of kernels
that score as weakly pigmented (highly paramutated) appear on the left
half of each histogram; more fully pigmented kernels are positioned in
the score categories to the right half of each histogram. The seedling
was raised in 22 C continuous light (LL) conditions for 15 days then shifted
to 12 hr light-dark cycles (LD) for six days before removal to field conditions
for testcrossing at maturity.
The pollen collections from the first three days that pollen was shed show most kernels scored in the lower half of the scoring range (fewer pigmented cells, most paramutation). The last three histograms show most kernel pigment scores in the upper half of the scoring range (least paramutation). When all eight histograms are compared, sequentially, the transition from frequencies of least to most pigmented kernels is visible as a gradient over the eight days of anthesis.
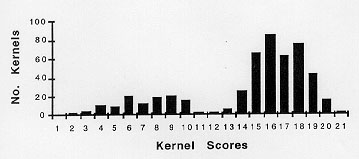
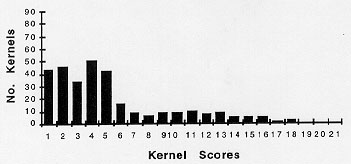
The pollen collections of Figure 1 represent epigenetic responses to controlled environmental conditions applied at a specific stage of development. More extreme changes in paramutation of R allele expression in response to environmental conditions can be seen in Figures 2 and 3. Kernel pigment-score profiles are skewed to the right and left halves of the histograms, respectively. All plants tested in Figures 2 and 3 were started in LL conditions at 22 C. Figure 2 represents testcross scores of kernels from eight plants that as seedlings received 12 hr LD cycles at 22 C for days 16-21. The darkest testcross pigment scores (least paramutation) were found in the last pollinations of single plants. Figure 3 shows pigment scores for testcrosses from the lowest tassel branches of seven plants that as seedlings were subjected to LL conditions at 32 C, days 15-21. These pigment scores are skewed to the left half of the histogram (more paramutation).
Figure 1 shows that different levels of epigenetic programming of R allele expression can be directed selectively at the earliest gametes from the upper part of the tassel or at the last gametes shed from the lower branches of the tassel. Figures 2 and 3 show that under different controlled conditions more or less paramutation can be programmed into the gametes from the upper or lower part of the inflorescence. The differences in gamete expression, under paramutagenic conditions, are the product of environmental conditions administered to meristematic tissue at a specific stage of development, the period during which the plant is susceptible to floral induction. Specific environmental conditions applied at the time of change-over from vegetative to floral stages of development identifies a threshold that makes it possible to regard the differences in the level of paramutant R allele expression as a useful reporter system for the more significant epigenetic event, that of floral induction. Significant differences in the number of nodes, tassel branches and time to anthesis (MNL 66) correlate with the differences reported as paramutation scores in the aleurone layer of the seed. Is it possible the r locus, responsible for a transcriptional activator, may play a more significant role in development than just the control of kernel pigment?
Since paramutation has been reported to be associated with methylation, the above information suggests a stage during development when methylation at the r locus can be initiated. The induction of change from vegetative to floral development is methylation associated in a species of Arabidopsis (Burn et al., Proc. Nat. Acad. Sci. 90:287-291). Light and temperature signals that time plant development in transgenic petunias are found to be highly correlated with methylation changes (P. Meyer et al., Mol. Gen. Genet. 243:390-399). Cause and effect relationships have been difficult to resolve in methylation studies. The epigenetic programming of the r locus as well as transgenes in petunia suggest that control of light and temperature conditions during early development will be essential if methylation is to be resolved as cause or effect when associated with gene programming.
Figure 1. Testcross kernel scores from a single plant showing the frequency of weakly pigmented and strongly pigmented kernels in testcrosses made each day over the eight days that pollen was shed. 50 pigmented kernels from each testcross ear were scored against a set of 21 standard kernels ranging from colorless to fully pigmented, 1 to 21 respectively. With each day that pollen was tested, frequency histogram profiles are gradually skewed to the right as anthesis progresses down the tassel.
Figure 2. Testcross scores of 50 kernels from each of eight plants which as seedlings received four LD cycles at 22 C followed by two LL cycles at 32 C, days 16-21. Up to day 16, seedlings were held in 22 C LL conditions.
Figure
3. Testcross scores of 50 kernels from each of seven plants which as
seedlings received seven 32 C LL cycles, days 15-21. Up to day 15, seedlings
were held in 22 C LL conditions.
Return to the MNL 70 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}