Embryo salt soluble proteins as markers in research on the biological
background of heterotic gene expression
--Drinic, SM; Coric, T and Konstantinov, K
A better understanding of the biochemical basis of heterotic gene expression could enhance the breeder's ability to form new maize genotypes expressing "permanent" heterosis. Several genetic models for the explanation of hybrid vigor in Mendelian terms have been suggested, including the dominance and overdominance hypotheses. The dominance hypothesis attributes the increased vigor of heterozygosity to dominant alleles and in principle should be fixable by inbreeding. The overdominance hypothesis assumes that there exist relatively rare loci at which the heterozygote is superior to either homozygote but heterosis due to overdominance or pseudo-overdominance and is not fixable by inbreeding. There are also results providing clear evidence for the role of epistasis (Russell and Eberhart, Crop Sci 10:165-169, 1970), and also indications that additive genetic effects are primarily responsible for the increase of heterosis through 5 cycles of maize selection and population crosses (Walejko and Russell, Crop Sci 17:647-651, 1977).
Maize hybrid plants expressing heterotic vigor develop from embryos consisting of 2n chromosomes which have not changed by the process of genetic recombination. Parameters derived from hybrid embryo genome expression could provide more information on the relationships between genome expression of parental lines per se and hybrid genome expression as a consequence of inbred line combinations. Salt soluble proteins, fractions of metabolically active albumin and globulin proteins, are good candidates for such studies.
A diallel set of five inbred lines, F2, ZPL120, ZPL203, W401 and EP1, excluding reciprocal crosses, has been studied at two locations, in a random block system experiment with 4 replications. Results on the heterotic effect on grain yield and soluble protein content of 10 developed F1 hybrids are presented in Table 1.
Table 1. Heterotic effect on the grain yield and soluble protein content
in the embryo of all developed hybrid combinations, and index of similarity
of inbred lines.
|
|
|||
| Hybrid combination | grain yield | salt soluble proteins 10-3 g/g fresh weight | index of similarity |
| ZPL120 x W401 | 139.43** | 8.28 | 79.2 |
| W401 x EP1 | 134.30** | 4.14 | 83.3 |
| ZPL120 x EP1 | 110.52** | 6.44 | 85.7 |
| F2 x ZPL120 | 109.39** | 3.44 | 85.2 |
| ZPL120 x ZPL203 | 90.97** | 4.52 | 86.3 |
| ZPL203 x EP1 | 90.34** | 0.96 | 85.2 |
| ZPL203 x W401 | 88.48** | 3.34 | 86.8 |
| F2 x EP1 | 87.36** | 0.65 | 87.7 |
| F2 x W401 | 67.22** | -0.63 | 88.5 |
| F2 x ZPL203 | 51.96** | 0.42 | 88.0 |
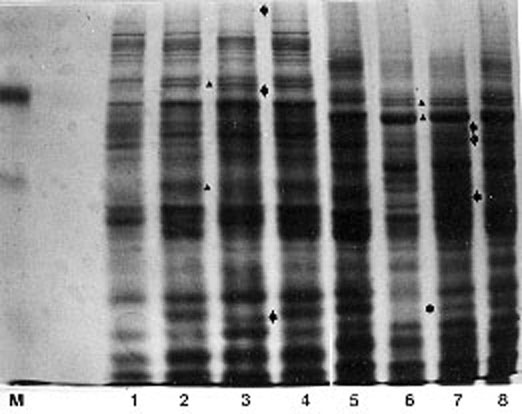
In all hybrid combinations significant positive heterosis was obtained for grain yield. Low but positive heterotic effect on the salt soluble protein content in embryo tissue has been obtained in 9 out of 10 hybrids. In order to correlate grain yield heterosis and salt soluble proteins in embryo tissue several hybrid combinations were selected for high resolution polyacrylamide gel electrophoresis (PAGE) of embryo salt soluble proteins: two hybrids expressing high heterosis for grain yield and two hybrids expressing low heterosis for grain yield. In both groups, hybrids have one inbred line as a common parent. Electrophoregrams are presented in Figures 1 and 2, respectively. In the hybrid combinations expressing the highest heterosis for grain yield and salt soluble protein content (F2 x ZPL120; ZPL120 x W401) 3 hybrid-specific protein bands are identified (arrows in Fig. 1). Several male or female inbred-specific protein bands have also been identified amongst the many in common for both parents.
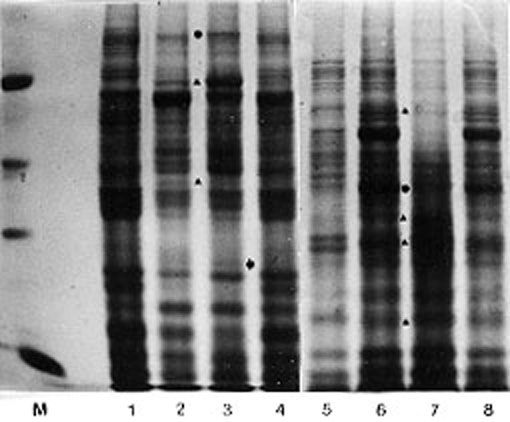
On the electrophoregrams of salt soluble proteins isolated from the embryo tissue of low heterotic hybrid combinations, presented in Figure 2, only one hybrid specific protein band has been identified (arrow).
By comparing electrophoregrams of hybrid combinations (F2 x ZPL120 and ZPL120 x W401; F2 x ZPL203 and F2 x W401) and coelectrophoregrams (F2 + ZPL120 and ZPL120 + W401; F2 + ZPL203 and F2 + W401) of parental lines it could be suggested that non-additive genetic effects are responsible for salt soluble protein synthesis in hybrid embryo tissue (Leonardy et al., TAG 82:552-560, 1992).
Based on the number of protein bands, their distribution according to the molecular weight, and presence or absence, the index of similarity of parent lines in hybrid combinations has been calculated and is presented in Table 1. Comparing heterosis for grain yield and protein content with index of similarity of inbred lines it could be suggested that inbreds with the lowest level of salt soluble protein similarity expressed the highest heterosis both for grain yield and content of salt soluble protein in embryo tissue.
The above data indicate that there is hybrid specific expression of certain loci in maize embryo tissue, and further biochemical experiments (i.e. isolation of poly A-mRNA specific for the protein bands synthesized only in hybrid combinations) for a better understanding of the molecular basis of heterosis are in progress.
Figure 1. Electrophoregrams of embryo salt soluble proteins of two hybrids expressing high heterosis for grain yield: line 1 (F2); line 2 (ZPL120); line 3 (F2 x ZPL120); line 4 (F2 + ZPL120); line 5 (ZPL120); line 6 (W401); line 7 (ZPL120 x W401); line 8 (ZPL120 + W401).
Figure
2. Electrophoregrams of embryo salt soluble proteins of two hybrids
expressing low heterosis for grain yield: line 1 (F2); line 2 (ZPL203);
line 3 (F2 x ZPL203); line 4 (F2 + ZPL203); line 5 (F2); line 6 (W401);
line 7 (F2 x W401); line 8 (F2 + W401).
Return to the MNL 70 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}