Plants homozygous for the iojap (ij) mutation have patterned white stripes on their leaves (Jenkins, J.Hered. 15:467-472, 1924), and give rise to albino seedlings in their maternal progeny when pollinated by wild-type plants (Rhoades, Cold Spring Harbor Symp. 11:202-207, 1946). The degree and extent of striping, and the maternal transmission of the ij-affected plastids, are largely dependent on genetic background (Jenkins, 1924; Coe, Thompson and Walbot, Am. J. Bot 75:634-644, 1988). The Ij gene was cloned by transposon tagging, but the sequence of the IJ protein showed no homology with proteins in current databases (Han , Coe and Martienssen, EMBO J. 11:4037-4046, 1992). To further characterize the role of the IJ protein in chloroplast and leaf development, antibodies were raised against recombinant IJ fusion proteins. Western blotting and immunolocalization have revealed that IJ is a chloroplast protein found in most cell types of immature leaves (not shown).
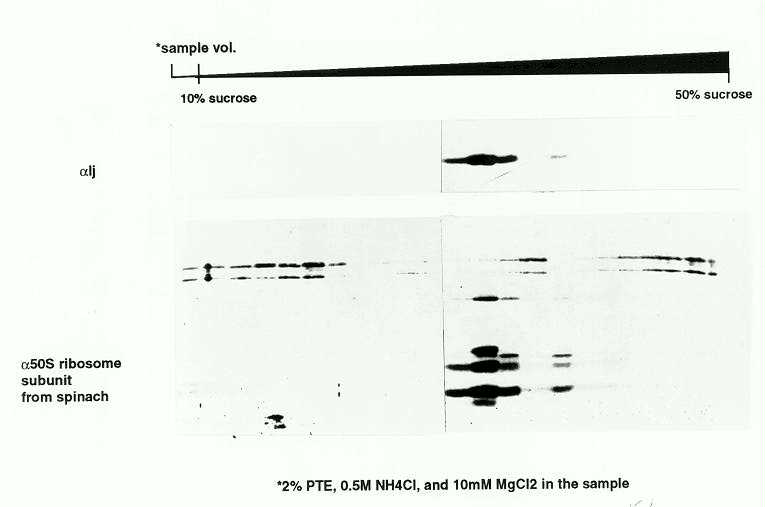
On the basis of ribosomal RNA profiles of mutant and wild-type plants, Walbot and Coe (Proc. Natl. Acad. Sci. 76:2760-2764, 1979) proposed that iojap plants suffered a programmed loss of plastid ribosomes that could account for the maternal transmission of ij-affected plastids. We therefore analyzed plastid subfractions to determine whether IJ was associated with plastid ribosomes or other macromolecular structures. In chloroplast lysates, most of the protein was found in high speed pellets, but it was found in the soluble fraction in the presence of 20mM EDTA. This suggested that the IJ protein was associated with an EDTA-sensitive complex. Whole seedling lysates were further fractionated by pelleting through a sucrose cushion, followed by sucrose gradient sedimentation in the presence of non-ionic detergents (polysome gradients). Sedimentation profiles were determined by uv absorption (not shown) and by western blotting (Fig. 1) using anti-IJ and anti-50S ribosome antibodies (from R. Mache). The IJ protein was found in the same fractions as the 50S ribosomal proteins under these conditions (Fig. 1).
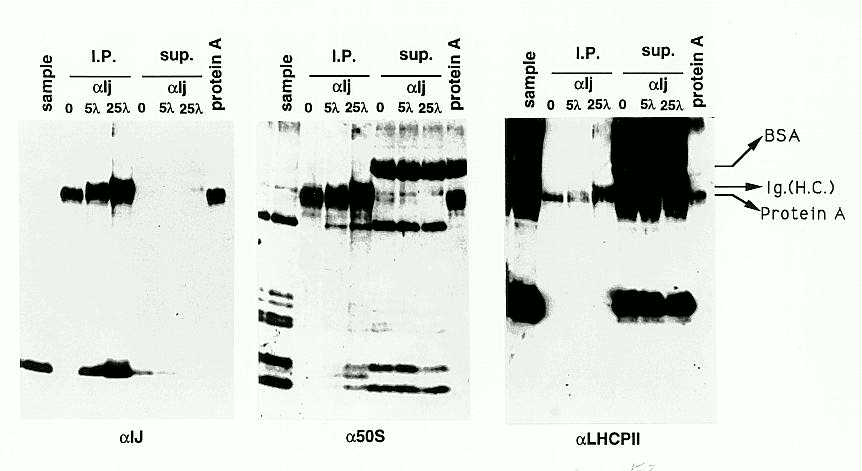
Direct association of IJ with 50S subunits was examined by immunoprecipitation with affinity purified anti-IJ antibodies. Immunoprecipitates were fractionated by SDS PAGE and western blots were probed sequentially with anti-IJ, anti-50S ribosomal proteins and control anti-LHCPII (light-harvesting complex membrane protein) antisera (Figure 2). IJ protein was quantitatively precipitated by increasing amounts of anti-IJ antibody (first panel). 50S ribosomal proteins were also specifically recovered in the anti-IJ immuno-precipitates (second panel, I.P.), although most of the proteins remained in solution (second panel sup.). LHCPII proteins were not recovered in the precipitates (third panel), demonstrating that the co-immunoprecipitation was specific, and did not reflect contamination with residual membrane-bound ribosomes. We are currently determining which ribosomal proteins are specifically bound to IJ.
Association with 50S ribosomal subunits suggests a role for the IJ protein in the regulation of plastid translation, and supports the model first proposed by Walbot and Coe for the programmed loss of plastid ribosomes in ij mutants: Some ribosomal proteins are encoded by plastid genes, so that loss of plastid translation due to homozygous nuclear mutation could lead to the loss of plastid-encoded ribosomal proteins. Restoration of the nuclear components of the plastid translational apparatus in heterozygous maternal progeny would not be expected to restore plastid ribosomal function if the plastid-encoded components were no longer present. Heritable ribosome-deficient plastids would result. Consistent with this model, IJ protein is severely reduced in abundance in defective plastids inherited cytoplasmically from variegated plants (not shown).
Figure 1. Western blots of IJ and 50S ribosomal proteins fractionated by sucrose gradient sedimentation. Plastid lysates were pelleted through 0.75M sucrose cushions, resuspended in 0.5M NH4Cl and 2% PTE, and were loaded onto a 10-50% sucrose gradient. The gradient was centrifuged at 150,000g for 16hrs, and fractions were collected. Proteins from each fraction were analyzed by SDS-PAGE, and by Western blotting using either affinity purified anti-IJ antibody (top panel) or anti-spinach 50S plastid ribosomal protein antibodies (lower panel; from R. Mache) respectively.
Figure
2. IP-westerns using anti-IJ antibodies. Samples were pelleted through
sucrose cushions (Figure 1) were resuspended in 50mM Tris pH8, 5mM MgCl2,
20mM KCl, and were used for immuno-precipitation with affinity-purified
antibodies. The immuno-precipitates and supernatants were analyzed to detect
IJ protein, 50S ribosomal proteins, and LHCPII membrane proteins, respectively,
by sequential probing of the same blot. Lane 1, sample before i.p.; Lanes
2, 3 and 4, i.p. with 0, 5 and 25 microlitres of anti-IJ antibody; Lanes
5, 6, and 7, supernatants from immuno-precipitates loaded in lanes 2, 3
and 4, diluted 5-fold relative to the precipitates. The last lane was loaded
with protein A beads alone.
Return to the MNL 69 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}