Organogenesis of the maize mutant Fascicled ear (Fas)
--Gretchen Haas and Alan Orr
Fas is one of the earliest acting mutants on the development of the inflorescence axis of maize. The pronounced, normal elongation of the apical meristem that occurs upon transition from the vegetative state to the reproductive state is disrupted by Fas (Postlethwait and Nelson, MNL 64:81-82, 1990). Key abnormal organogenic features include a shift in the direction of transition meristem growth (width doubles its height) and a bifurcation of the broad transition meristem into two primary inflorescence axes, each with a terminal apical meristem. This bifurcation of each terminal apical meristem may be repeated several times in the ear and central spike of the tassel before the process ceases. Since the appearance of terminal apical meristems of Fas inflorescences is similar to the spikelet pair (branch) primordia before the formation of pedicellate and sessile spikelet pairs, it was suggested by Hessler, R (1963) and Postlethwait and Nelson (1990) that Fas "time-shifts" the bifurcation event to an earlier ontogenetic stage.
There is no previous SEM organogenic study of this mutant, although an examination of Ruth Hessler's dissertation reveals a histological description of Fas ear and tassel development. We undertook this investigation to establish a morphological series of development stages for sampling stage-associated proteins in Fas inflorescences. This permits us to test, at the molecular level, the 'time-shift hypothesis': whether a subset of putative protein markers of normal spikelet pair primordia (Coffe, Findlay, Wagner and Orr, Int. J. Plant Sci. 153:31-39, 1992) is found in the terminal meristem of mutant inflorescences.
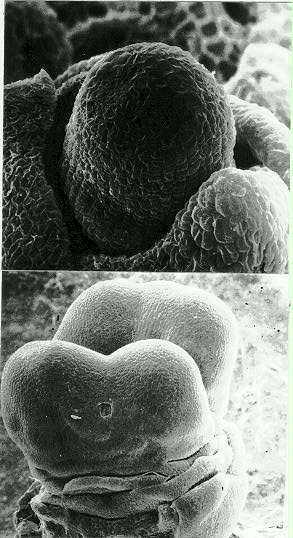
An SEM examination of more than 40 developing ears and tassels of Fascicled ear confirmed that, in the majority of cases, mutant organogenesis followed the pattern described in Hessler's histological studies. Figure 1 illustrates an ear inflorescence whose apical meristem is at the transition stage. This enlarged width stage is followed by a bifurcation of the apical meristem. Figure 2 shows an ear where a second bifurcation cycle has been initiated. Initiation and early development of spikelet pair, spikelet, and floret primordia on each derived Fas inflorescence axis is essentially the same as expected for normal maize ear and tassel development, except the inner surface of each axis is often barren of primordia.
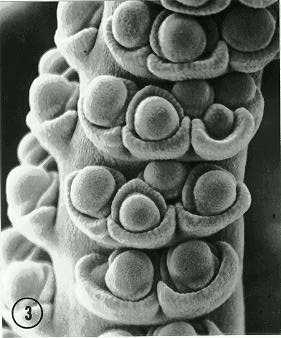
However, based on these SEM studies of ear and tassel development in Fas we found additional alterations to the normal sequence of inflorescence organogenesis. In several instances 1-2 additional ranks of spikelets were produced along the clustered, bifurcated ear axes (Fig. 3) and the bifurcated central spike of the tassel. Figure 3 illustrates, at the base of the photo, the usual paired arrangement of pedicellate and sessile spikelets (each with a lower floret initial); note the additional spikelets at the two nodes proximal to the lower paired spikelets. In these cases the extra spikelets were smaller and at an earlier stage of development. If the execution of the mutant bifurcation program is pronged into the spikelet pair stage, it appears the Fas gene occasionally is expressed at the switch point between spikelet pair primordia and spikelet primordia. This is similar to a second round of bifurcations in spikelet pair primordia of Argentine popcorn (Sundberg and Doebley, JF, MNL 64:21-22; Sundberg and Orr, unpublished), where a doubling of the row number shifts the popcorn inflorescence from distichy to polystichy.
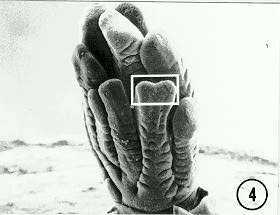
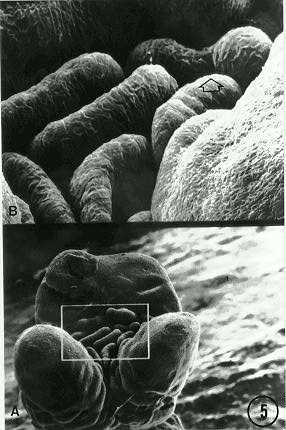
We also observed in tassels a previously unreported bifurcation of the distal tips of the secondary (lateral) branches (Fig. 4, box). It seems the mutant program is retained in the branch primordia that give rise to the elongate lateral branches of the tassel. This also is consistent with a prolonged expression (bifurcation) of the Fas gene at the spikelet-pair-primordia switch point. Another new observation is the presence of numerous bract primordia within the ring, and at the base, of a cluster of mutant derived female inflorescence axes (Fig. 5). Note in Figure 5 (arrow) the bifurcation of one bract primordium. We are unclear whether these observations represent an expression of the Fas gene in the vegetative phase.
We are currently testing a time displacement model for a better understanding of the regulatory events in maize floral development. This is based on an analysis of 2-D PAGE protein extracts from selected inflorescence primordia of Fas.
Figures 1-2. Early ear organogenesis. 1. Ear at transition stage of development. The specimen shows a broad shoot apical meristem prior to the first bifurcation at the distal tip. 2. A second bifurcation of each new inflorescence axis is shown.
Figure 3. Single inflorescence axis from a Fascicled ear cluster of female inflorescence after several bifurcation cycles. Inflorescence is characterized by extra spikelets at the three upper nodes. The lower node shows paired spikelets with glumes, lemmas and lower floret primordia.
Figure 4. Fas tassel after a few bifurcations of central spike tissue. Tips of long lateral branches are bifurcated (box).
Figure
5. Female inflorescence of Fas with numerous bracts clustered
at base (A, box) of several inflorescence axes derived from bifurcations
of the terminal meristem. Note bifurcation of a single bract (B, arrow).
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}