--G. Borovsky and V. Vojnikov
It is known that the maize cell under heat stress synthesizes heat shock proteins (HSP) having a protection function. It is assumed that some HSP can prevent denaturation of proteins thus increasing resistance of the cell to heat. According to this hypothesis we performed the experiments with glucose-6-phosphate dehydrogenase (G6P-DH) in vitro. We aimed to find the effect of plant HSPs on the activity of G6P-DH after heating. The denaturation of G6P-DH was inferred from the decrease of its activity.
Proteins were extracted from 3-d-old etiolated seedlings of maize (Vojnikov, VK et al., Plant Physiol. 33:221-225, 1986). Seedlings were grown at 27 C (CP), or heat shocked during the last 16h at 41 C (SP). We used the whole water soluble fraction of proteins precipitated by 4 volumes of cold acetone. Proteins were solubilised in 0.1M Tris-HCl buffer and were frozen in liquid nitrogen in small portions. Each portion was used immediately after melting. Bovine serum albumin (BSA) was prepared by the same procedure. G6P-DH activity was measured according to Kornberg et al. (Methods Enzymology, NY, 1955) using test-combination (Boehringer Mannheim). G6P-DH (Serva) was used in 0.000125mg/ml concentration and prepared immediately before the experiment. Heating was carried out in buffer from test-combination. G6P-DH solution was incubated for 10 min with (or without) maize proteins or BSA at 25, 35, 45 C. Then the temperature of solution was rapidly (1 min) reduced to 25 C and the activity of G6P-DH was measured. Statistical calculations were made by standard methods. The activity of G6P-DH after heating was determined in percentage from the activity at 25 C. The activity of proteins from maize was subtracted from the total. It was less than 8% of the total activity under maximal concentration (100mg/ml) of maize proteins in the mixture.
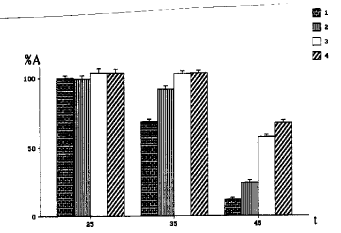
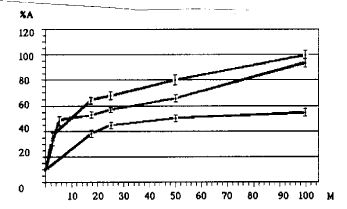
No decrease of the activity of G6P-DH and no effect of CP, SP and BSA on this activity were found after 10 min incubation at 25 C with the particular concentration of plant proteins (25mg/ml). After incubation at 35 C without any proteins, the activity of G6P-DH was 69%; after incubation with BSA - 92%; and after incubation with CP or SP - 103-104%. Following incubation at 45 C without any other proteins, the activity of G6P-DH was 10%; after incubation with BSA - 24%; with CP - 57%; with SP - 68% (Fig. 1). Using different amounts of maize proteins and BSA the percent of protected activity of G6P-DH is found to increase to 50% with increasing added protein concentration to 12.5mg/ml, and to almost 100% of the protected activity with increasing added protein concentration to 100mg/ml after incubation at 45 C (Fig. 2). The activity of G6P-DH after incubation with SP was higher than with CP by 10-12% at all concentrations except for the activity of G6P-DH close to 0 or 100% (Fig. 2).
A factor has been found among maize proteins which protects the activity of G6P-DH in vitro at high temperatures. This factor is assumed to be heat shock proteins (HSPs) and heat shock related proteins (HSRPs). The action of CP and SP is similar and the differences are probably related to the content of active proteins in total fraction or to the ability of the proteins with similar functions to effect protection. The synthesis of HSPs and HSRPs is known to be necessary for the cell to be alive at all temperatures (Ang, G and Georgopoulos, C, J. Bacteriol. 171:2748-2755, 1989; Werner-Washburne, M et al., Mol. Cell. Biol. 7:2568-2577, 1987) therefore, we believe that HSPs and HSRPs are present in our fraction of maize proteins.
It appears that some HSPs and HSRPs function on the molecular level as chaperones or molecular conformation designers (Beckmann, R et al., Science 248:850-857, 1990; Ostermann, J et al., Nature 341:125-130, 1989). The fraction of proteins precipitated only by high concentrations of ammonium sulfate serves as an anti-denaturation factor affecting proteins in vitro. It is interesting that this fraction was enriched by low molecular weight HSPs (Jinn, T et al., Plant Cell Physiol. 30:463-471, 1989). These facts are consistent with our hypothesis that HSPs play the main role in thermoprotection in our experiments.
The mechanisms of stabilization of proteins are unknown. On the basis of the main principles of protein denaturation (Jaenicke, R, Prog. Biophys. Molec. Biol. 49:117-237, 1987) we assumed that the anti-denaturation factor is connected with macromolecule at the points important for its flexibility, thus increasing the rigidity and thermostability of the molecule.
Figure 1. Activity of glucose-6-phosphate dehydrogenase after heating at different temperatures. The standard deviation is shown. % A - activity after heating; t - temperature; 1 - G6P-DH without any other proteins; 2 - G6P-DH + BSA; 3 - G6P-DH + CP; 4 - G6P-DH + SP.
Figure
2. Dependence of activity of glucose-6-phosphate dehydrogenase after
heating on concentration of added proteins. The standard deviation is shown.
% A - activity after heating; M - amount of added proteins (mg/ml); upper
curve - G6P + SP; medium curve - G6P-DH + CP; low curve - G6P-DH + BSA.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}