DEFIANCE, OHIO
Defiance College
Paramutation: A system for temperature programmable transgenerational genetic change coupled to tassel determination
--Bernard C. Mikula
Paramutation, as reported by the late R. A. Brink, RA in 1956, involved the production of a heritable change in the kernel pigment expression of the R gene by its allele R-st. When testcrossed, all R genes emerging from the RR-st heterozygote were changed. Effort has been directed toward achieving binary control of paramutated R gene expression by controlling early developmental conditions of seedlings with a paramutation sensitive R gene. In Vol. 65 of MNL, I reported that, in testcrosses, near binary control of the level of paramutation of the R gene was achieved in plants which as seedlings were grown for their first three weeks under 28 C or 22 C temperatures. Sib seedlings of the 1990 RR-lst heterozygotes were used in 1991 to explore the developmental time interval when paramutation could be subject to temperature control. Seedlings were grown in constant light and temperature conditions until ready for floral induction. Light was provided by 14 VHO 200W daylight type fluorescent lights supplemented by 12 60W incandescent lamps. Seedlings were started 60cm from the light source and grew toward the light for the period of time held in the growth chamber. By day 10 seedlings raised in constant light at 32 C showed necrotic conditions in the youngest leaf tissues, a typical calcium deficiency symptom even though adequate calcium was present. This necrosis in the youngest leaves takes place when seedlings are ready for floral induction. Four L:D cycles (12h light:12h dark) relieved these symptoms and induced tassel formation. Floral primordia, however, were not visible until over a week later when seedlings were transplanted to field conditions. At the lower temperature, 22 C, tassel induction must take place later; six L:D cycles were applied to seedlings beginning the 16th day. At the end of each of the L:D treatment-periods plants were removed to the field to relieve crowding in limited growth chamber space. At maturity, those plants, raised as seedlings in controlled temperatures, were testcrossed to W22/W23 colorless recessives to assess the level of pigmentation of the R gene resulting from paramutation by the R-lst allele at the two different temperatures applied to the seedlings in their first two or three weeks of development. The R gene used was one of four new accessions from Native Seeds/SEARCH which showed greater responsiveness to temperature than a highly inbred R allele long maintained in W22 background. The R-lst allele was chosen because of low pigmentation in the aleurone, making it possible to assess the level of paramutation in the R gene by visual inspection of testcross ears. Because of space limitations, only eight seedlings for each different test could be used since crowding becomes an important factor toward the end of the two or three-week period.
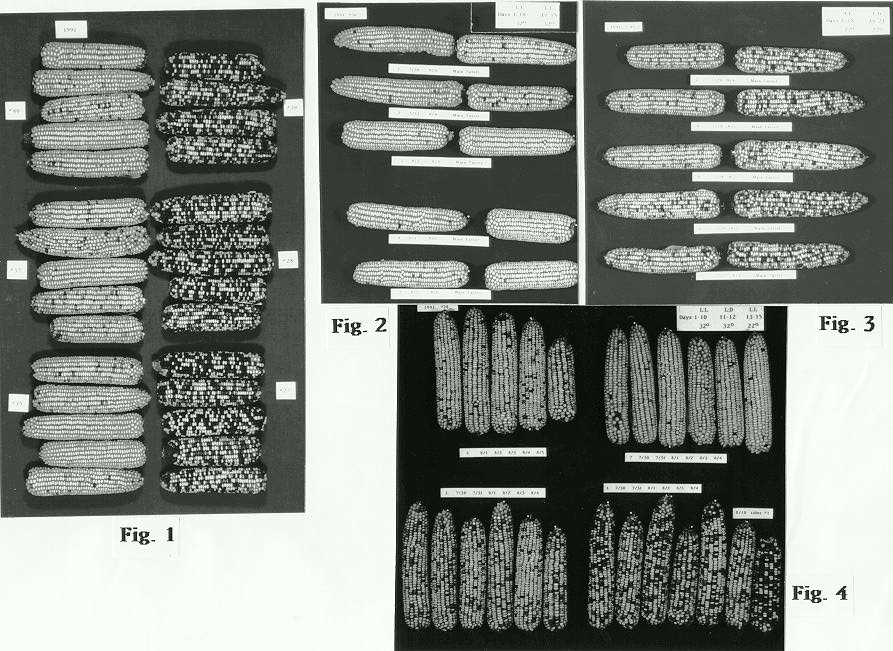
In Vol. 65 of MNL the photographs of testcross ears of paramutated R genes from 1990 plants whose seedlings were grown at 28 C showed a high level of paramutation; that is, the kernels were nearly colorless. These highly paramutated R genes, when tested for heritability, were found to remain considerably lighter (left column of Fig. 1) than R genes from plants whose seedlings received the 22 C conditions for their first 21 days (right column of Fig. 1). In 1991, an effort was made to define a shorter developmental time interval when paramutation could be experimentally controlled. Fig. 2 shows that seedlings which received 32 C and constant light for their first 15 days have paramutant R expressions, when testcrossed at maturity, approaching the colorless condition as reported for the previous year. Sib seedlings grown at 22 C through 21 days, with L:D treatments for days 16-21, showed testcrosses whose R gene pigmentation was much more darkly mottled (Fig. 3).
Plants grown as seedlings at 22 C (Fig. 3) for the three week period showed a tassel mosaic when testcrosses from a single tassel were compared over the seven day period that pollen from a single plant is shed. The two columns of testcross ears in Figures 2 and 3 represent the first and last pollen collections from each plant. Only the first pollen collections from seedlings which received 22 C show paramutant R expressions in testcrosses visibly lighter than those made later in the seven-day period (Fig. 3). A total of 25 plants of the 29 plants tested which received four (days 16-19) or six (days 16-21) L:D treatments at 22 C show the tassel mosaic when earliest pollinations from a single tassel are compared with the latest pollinations.
A high level of paramutation was achieved if seedlings were held at 32 C through the 15th day. By comparison, plants maturing from seedlings which received the lower temperature of 22 C show less paramutation has taken place. These two opposing degrees of paramutation can be used as limits for assessing results of further experiments designed to define more closely the developmentally sensitive period when seedlings respond to temperature treatment. Earlier results had shown that tassel determination for inbred W22 required a minimal four-day period of L:D treatments beginning day 10 at temperatures of 25 C. At 22 C tassel determination requires a six-day period of L:D beginning day 16. Figure 4 shows all the testcrosses from four different plants whose seedlings were held in constant light for ten days at 32 C then subjected to a two day L:D period of 32 C before transfer to 22 C. A greater range of variation in paramutation from plant to plant was observed (Fig. 4) than from those seedlings held for the same period in continuous light at 32 C (Fig. 2). A variety of levels of paramutation result if two-day temperature changes are inserted during this induction period regardless of whether plants were started at the lower or higher temperature regime (Fig. 4). It may be inferred that some plants, because of differences in developmental timing, escape the brief temperature interruption and are determined to be already highly paramutated; others, in response to the brief low temperature of 22 C, will retain a low level of paramutation. Three different experiments involving such inserted two-day temperature perturbations at this floral initiation period showed that a sensitive stage in development has been identified when temperature can make a significant input into the level of plant to plant variation for paramutant R gene expression. The mosaicism attributed to paramutation can now be considered influenced by temperature at a critical stage of development defined by the period plants are susceptible to floral induction.
Because induction of flowering requires the imposition of a
series of dark periods at different seedling ages, depending on
the temperature, it was thought the light:dark (L:D) cycles were
responsible for control of the level of paramutation. R
gene expressions from plants which received constant light were
lighter than those which received L:D periods. However, with
higher temperatures, the greater reduction in the level of R
expression suggested that temperature may be the more important
variable. As
seedlings grow increasingly closer to the radiant energy source in the closed system of a growth chamber, the application of dark periods results in lower plant leaf surface temperatures. From this it can be inferred that in constant light, temperatures are uniformly higher during the critical induction period hence the higher level of paramutation.
The results above show that near binary control of R gene expression under paramutagenic conditions can be achieved by manipulation of temperatures at a critical stage of development. A continuum of expression (heterogeneity) has been reported by the Wisconsin laboratory for the paramutant R alleles raised under field conditions. If paramutation can be used as a model, experimental operations describing the behavior of controlling elements in two-element systems under field conditions will require more complex operational description. The work at Wisconsin over the past 40 years, as Brink, RA pointed out in the early 60's, shows that allelic history must be taken into account in descriptions of paramutant R behavior. Allelic associations in previous generations and backgrounds can influence future behavior of the R gene quantitatively (number of cells in the aleurone layer). Efforts designed to explore the behavior of R genes under paramutagenic conditions using inbred backgrounds exclusively, can now be questioned. As we try to understand paramutation can we assume the gene will behave more "appropriately" in an inbred background without some test? What test? Can we be sure the paramutant R has not undergone a significant change in its potential behavior in the backcross process? Of the many different alleles available, which would be most useful for a chosen experimental design? In the backcrossing process which phenotypic expressions should be chosen for the continuation of the backcrosses? If temperatures have a significant effect early in development, can we rely on field-grown material in the future, since spring field conditions are notoriously variable from day to day? Another common assumption is that gene expressions from the same tassel are uniform, but the tassel mosaics under temperature control mean that several pollinations must be made on different days to be sure of the range of variation of the R gene from a single plant. To make operational description even more complex, the paramutant R can add the changes in R expression resulting from the presence of each of two paramutagenic alleles (RR-st R-st trisomics) in a single generation, or from year to year if exposed successively to paramutagenic alleles. These many questions, though distressing in the light of Mendelian canons, are the necessary prelude to the discovery of experimental operations for dealing with controlling element (transcriptional activator?) behavior at critical developmental stages. The experimental operations and assumptions supporting the Mendelian paradigm can no longer be a reliable guide for systems showing the characteristics of the paramutation-like systems. Though evidence is still lacking for an element at the R locus, both McClintock, B and Brink, RA considered paramutation an example of a two-element system.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}