Cold Spring Harbor Laboratory
COLUMBIA, MISSOURI
USDA-ARS and University of Missouri
Molecular cloning of ij (iojap): insertion of a Ds-like element is responsible for the ij-ref mutation
--Chang-deok Han, Robert A. Martienssen and Edward H. Coe
To understand the diverse and unique phenotypic expression of the iojap mutant, a transposon tagging experiment has been conducted using Robertson's Mutator (Mu). A new ij mutant was obtained from crosses of ij-ref (the original reference allele) to Mu lines. Subsequent genetic and molecular studies showed that the mutant was of a new ij allele (ij-mum1) from the Mu lines and contained a Mu1 element that cosegregated with the iojap phenotype (MNL 63:65 and 64:46).
A 6.0kb EcoRI genomic DNA fragment containing the Mu1 element in ij-mum1 was cloned. Flanking DNA from the genomic clone detected a single copy fragment in the genomic DNA and a 1.3kb transcript in leaf mRNA. The patterns of transcripts hybridizing with flanking DNA from the genomic clone were correlated with the different ij genotypes. The level of transcripts from ij-ref/ij-ref was severely reduced and the size of the transcripts was slightly smaller than that from heterozygous and normal seedlings. The level of transcripts was about 50% reduced in heterozygous seedlings, compared with that in homozygous normal seedlings.
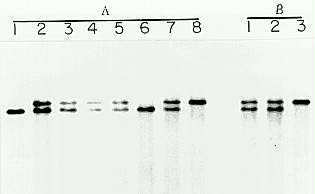
In order to verify that the cloned DNA was from the iojap locus, we took advantage of the fact that the original ij-ref allele described by Jenkins (1924) is unstable. Rarely, normal green sectors on pale green ('grainy') leaves have been observed in homozygous ij-ref plants even in an inbred background (Coe, EH et al., 1988). In 1979 and 1983, two independent germinal revertants (Ij-Rev) were obtained from plants in which green sectors ran through the tassel (Coe, EH, unpublished data). The Ij-Rev alleles were maintained by crossing to ij-ref/ij-ref plants and subsequent selfing. Normal plants (Ij-Rev/ij-ref or Ij-Rev/Ij-Rev) of selfed progeny of heterozygous Ij-Rev/ij-ref plants were examined by Southern hybridization, in comparison to ij plants carrying the ij-ref allele from the original plants with revertant sectors. Figure 1 is an autoradiogram of a Southern blot hybridized with a cloned DNA fragment flanking the Mu1 element. It shows that polymorphisms of BclI digested genomic DNA were correlated with the genotypes of the plants in two independent revertant families. Transcripts of the progeny of one of the germinal revertants were examined using the same probe (Fig. 2a). Northern blot data showed that transcripts in the revertant seedlings accumulated up to the normal level and were of the same size as those from normal siblings. Uniform loading of polyA+ RNAs in each lane was confirmed by reprobing the blot with maize actin genomic DNA as a probe (Fig. 2b). Also, eight independent somatic revertant sectors on the leaves of homozygous ij-ref plants were examined by Southern hybridization, in comparison to adjacent ij tissues in the same leaves. Two DNA fragments were detected in genomic DNA from all the green somatic revertant sectors. One of them was of the ij-ref size and the other one showed the same size as seen in the germinal revertant alleles (data not shown). The adjacent ij tissues contained one DNA band of the ij-ref size.
Figure 1. Southern analysis of the progeny of two independent germinal revertants (Ij-Rev). Genomic DNA was extracted from normal plants (Ij-Rev/ij-ref or Ij-Rev/Ij-Rev) from the selfed progeny of Ij-Rev/ij-ref plants of two families (A1-7 and B1-2) and from ij plants (lanes 8 and 3). The DNA samples were digested with BclI. The two independent revertants were designated A and B. The genotype of each normal plant was identified by progeny testing. Lanes A2, A3, A4, A5, A7, B1, and B2 were from heterozygous plants (Ij-Rev/ij-ref). Lanes A1 and A6 were from homozygous plants (Ij-Rev/Ij-Rev). Flanking DNA fragment of the genomic clone from ij-mum1 was used as a hybridization probe.
Figure 2. Northern analysis of transcripts from ij-ref, Ij-Rev, and normal (+/+) siblings. The blot of polyA+ RNA from ij-ref (lane 1), Ij-Rev (lane 2), and normal siblings (lane 3-5) was hybridized with a flanking DNA fragment of the ij-mum1 genomic clone (a). The following serial dilution of mRNA samples of normal siblings was loaded; 2 times higher than mRNA of ij-ref and Ij-Rev (lane 3), equal amount (4), 2 times dilution (lane 5), 5 times dilution (6). The amount of mRNAs in each lane was reexamined by reprobing the blot with a maize actin genomic DNA (b).
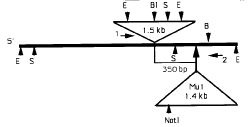
To understand the molecular basis of the reversion event at the iojap locus, we cloned a 6kb EcoRI DNA fragment from the ij-ref allele. Figure 3 is a composite restriction map of the genomic clones of ij-ref and ij-mum1. Except for 1.5kb additional DNA located around 350bp to the left of the Mu1 insertion site, the restriction map of ij-ref was identical to that of ij-mum1 except for the Mu1 element. Both ends of the 1.5kb DNA and the adjacent DNA were sequenced (Fig. 4a). The adjacent ij-ref DNA contains an 8bp direct duplication, relative to the ij-mum1 allele. And, both ends of the 1.5kb DNA have perfect inverted repeat sequences of 16bp. Eleven base pairs from both ends of the invert repeat are almost identical to the 11bp inverted repeats of Ac and known Ds elements. Terminal and subterminal regions of both sides of the element shared around 60% sequence homology with those of Ac and also have two or three DNA sequences that have been shown to be an Ac-transposase binding site (Kunze and Starlinger, EMBO J. 11:3177-3185, 1990).
Figure 3. A composite restriction map of the genomic clones of ij-ref and ij-mum1. The restriction maps of the ij-ref and ij-mum1 genomic clones were identical, except for an additional 1.5kb DNA and Mu1 element in each clone, respectively. An additional 1.5kb DNA of the genomic clone of ij-ref was located 300bp to the left of the Mu1 insertion site of ij-mum1. The 1.5kb DNA contained BglII, EcoRI, HindIII, and SstI sites. The locations of primers for PCR amplification are indicated as arrows (1 and 2) at the right of the 1.5kb of ij-ref and at the left of the Mu1 element of ij-mum1, respectively. Restriction enzyme sites are indicated as follows: B: BamHI, B1: BglII, E: EcoRI, H: HindIII, and S: SstI.
Figure 4A. Partial DNA sequences of the 1.5kb fragment inserted in the ij-ref allele. The sequences of 1.5kb DNA are in capitals, while the adjacent ij sequences are in lower case. The italic bases indicate the 8bp direct duplication of ij-ref. The 16 bases of bold character in both ends of the 1.5kb DNA show the inverted repeat. DNA sequences corresponding to the Ac transposase binding motif are underlined. Figure 4B. Terminal inverted sequences of the 1.5kb element of ij-ref, Ac and other Ds elements. Sequence differences in the 1.5kb element are underlined. The data are from Pohlman et al., 1983 for Ac, from Sutton et al., 1984 for Adh-Fm335, from Weck et al., 1984 for sh-m6233 from Wessler et al., 1986 for wx-m1.
To obtain molecular proof that functional and phenotypic recovery from the genetic lesion of ij-ref could be due to excision of the 1.5kb element, seven independent revertant alleles were analyzed for DNA sequences around the insertion site of the 1.5kb Ds-like element in ij-ref. Two germinal revertants described above and 5 new somatic revertant sectors were used. Comparison of partial sequences of the genomic clones and cDNA of Ij showed that the 1.5kb element was inserted into an exon of the gene. To clone DNA of the Ij-Rev alleles around the insertion site, DNA of Ij-Rev alleles was amplified by PCR (Polymerase Chain Reaction) using primers from the left of the 1.5kb DNA and from an intron right of the Mu1 insertion site. The PCR products were sequenced. Sequences outside the duplication region were identical in ij-ref and all 7 Ij-Rev alleles. Along with partial sequences of the genomic clones of ij-ref, ij-mum1 and cDNA of Ij, the sequences of the seven Ij-Rev alleles around the insertion site of ij-ref are shown in Figure 5. None of the PCR clones contained any sequences from the 1.5kb element. However, there were differences in the direct duplication sequences among the different alleles. Four different sequences were found from 7 independent revertants. One of the somatic revertants contained the same sequences as the wildtype, which indicated that the Ij-Rev allele was generated by perfect excision of the element. All the revertant alleles maintained coding regions in frame after the element was excised.
Figure 5. DNA sequences of Ij-Rev alleles. Genomic DNA from two germinal and 5 somatic revertants were amplified by PCR. The primers are indicated in Figure 3. Part of sequences of 7 Ij-Rev alleles are listed along with ij-ref, ij-mum1 and cDNA of ij. The duplicated host DNA is underlined.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}