--Mary Ann F. Cushman and Tim Helentjaris
Results from an earlier study (Edwards et al., 1991) demonstrated that the genomic locations of several regions (QTL) controlling quantitative variation for height in F2 progenies from the cross, CO159 X Tx303, coincided with the locations of a number of dwarf mutant loci. One particular QTL was closely linked by analysis of the isozyme, Acp1, and the anonymous RFLP, NPI222, to the centromere of chromosome 9 and to a GA-biosynthetic locus, defined by the mutation, d3. This region accounted for 27% of the variance for plant height in this particular experiment and was also an important yield QTL. Similar results have since been observed in a number of other crosses.
The following test was devised as part of a series of studies designed to test Robertson's hypothesis (J. Theor. Biol., 1985) that moderate alleles at loci previously defined by more extreme mutant alleles may be responsible for quantitative variation; that is, they may function as QTL. This experiment has been repeated three times on inbred maize lines, twice in the greenhouse and once in the field; the latest greenhouse experiment is described herein. Given positive results from these initial experiments, our intent is to apply this method to segregating populations to provide stronger supporting evidence for efforts to clone loci that may behave as important QTL.
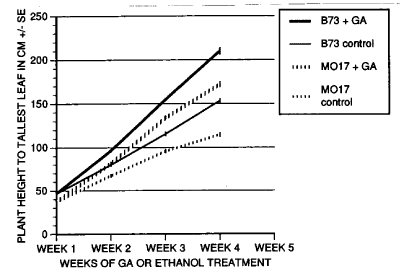
The inbreds Mo17 and B73 were planted in the greenhouse in August 1991 in a randomized complete block design with four replications and two blocks per replication. In each block, four plants of each genotype were treated weekly with either 100ul of 1mg/ml GA3 (0.1mg or 2.7X10-7 moles applied) in 40% ethanol or 100ul of 40% ethanol as a control. Height measurements for all plants were taken weekly, measuring from soil level to the tip of the longest leaf or to the top of the culm. At the end of the experiment, internode numbers were also recorded; however, this value was unaffected by GA treatment. Plant heights of the treated and untreated populations of each genotype differed significantly after a single treatment (Fig. 1, Week 2).
To describe the response to GA treatment, an index of growth was calculated for each plant in both the control (IC) and the treated (IT) populations, where I = (final height - initial height)/initial height. These genotypes differed significantly for the growth index when treated with ethanol alone in the control (Mo17 IC = 3.92±0.19 and B73 IC = 5.51±0.14). After four weeks of treatment with GA3, however, there was no significant difference in the growth index (Mo17 IT = 10.20±0.43 and B73 IT = 10.04±0.34), suggesting that these inbreds possessed the same potential for plant height once GA biosynthesis was no longer limiting. A simple measure of relative response to GA (RGA) might be calculated as RGA=IT-IC. These two genotypes would differ for this value (Mo17 RGA = 6.20 and B73 RGA = 4.53). It is possible that the difference in RGA for plant height in these two inbreds might be ascribed to simple differences in GA levels, presumably due to different alleles at GA biosynthetic or degradative loci. Height in progenies from this cross can be shown to vary quantitatively, so it will be important to show whether variance in a segregating population can be precisely explained by any of the GA biosynthetic loci. If similar results (IT values that are not significantly different, but RGA that are significantly different) are obtained with other inbred lines and segregating progenies, this may provide a system in which the limitation of GA biosynthesis can be evaluated as a QTL controlling plant height in various genetic backgrounds.
These results demonstrate that these lines differ significantly in height both with and without treatments with GA3, and for an index of growth in the absence of exogenously added GA, but not for the index of growth following GA treatment when GA is non-limiting. We conclude that it may be possible to partition the variance for plant height with and without exogenously added GA and demonstrate the presence of loci which quantitatively control endogenous GA levels and hence, plant height.
One way to address this question more precisely may be to use inbred
lines into which different alleles of a region which may contribute to
quantitative variation have been introgressed. These lines could be compared
to their corresponding recipient lines for height variables with and without
exogenously added GA. In this case, the importance of specific loci can
now be assessed more precisely, because the confounding effects of other
regions have been removed. In a sense, one has converted quantitative variation
into qualitative variation. By introgressing a single region presumed to
contain the d3 locus into a number of genetic backgrounds, one should
be able to compare the action of this new allele very precisely to the
original allele present in these backgrounds. These lines should be equivalent
in their ability to respond to GA, but may differ in endogenous GA levels,
presumably due to the action of different GA alleles. If the paired lines
(recipient and converted lines) differ in plant height without treatment
but do not differ significantly after GA treatment, this would be strongly
suggestive that the d3 locus was functioning as a major QTL for
plant height in these inbreds and would confirm a major component of Robertson's
hypothesis. It would also demonstrate that this could be a productive strategy
to confirm and clone genes that act as major QTLs for a number of important
agronomic traits.
Return to the MNL 66 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}