On the other hand, based on our first rainbow estimate of latente linkage, fl1 seemed to be one possible marker for it. By one cross and four backcrosses we obtained the floury-1 versions of 2 Tuxpan lines, 2 Cateto lines, 1 ETO flint line and 1 San Luis Potosi line, derived from the same source as in MNL 61:27-29. These floury versions were crossed to a latente source and used as recurrent parents. In this material we husked the tips of the ears. In the SLP backcrosses there was a segregation for color, so color acted as dominant. It appeared in all its progenies, which were plants selected by means of a Steady State Porometer Licor-1600 apparatus in the previous year. The plants were under severe atmospheric stress by heat and very low relative humidity. The data are presented in Table 1 including also kernel row numbers and the analysis. An unspecified linkage Krn Fl1 is readily visible comparing the factorials Krn x C with Fl x C (C = uncolored) but the data can not be explained by only one pair of alleles. In Fl C factorial one sees many more colored (39%) in the fl1 line than in the Fl1 (19%) and again the variation can not be explained by only one pair of alleles. The models which explain the data are presented in Table 1, giving nearly perfect fit for Krn Fl1 and a reasonable one for Fl1 C. The sum of the probabilities in each 2x2 factorial is one. To correct it to the population size n=87, we can multiply them by 87 or correct the calculated values in the end. In the case of Fl Krn for example, if Fl1 Krn2 (see MNL 61:27-29) linkage is q, and Fl Krn12 linkage is p, the joint probability of Fl1 bringing both together is (l-p)(l-q), and so on. Taking the logarithm likelihood expression including a, b, c and d, differentiating in relation to p and q, and multiplying by minus one to simplify we arrive at
and p=30.0+10 and q=32.2+10, the errors being calculated by Fisher's indirect method (see MNL 62:37-38), with survival equal to 1 for all entries, a very rough estimation, we guess. The inclusion of allometric coefficients did not improve the results.
For Fl1 and colour we did Fl1 ltp recombination value (ltp = latente p color) equal q and Fl1 p2 (p2 because it is similar to existent p1) as p, and p2 fl1 is p=30.0+10 and q=33.3+11.
In maize x teosinte studies kernel row numbers should be counted and this model applied.
The high frequency, at least in our reported work of these types of situations, suggests that in QTL x RFLP programmes these models should be included in the computer in order not to lose important results.
The systematic occurrence of flavone
glucoside factors linked to Krn suggests that these blocks may be
the triggers for a system of horizontal resistance to pests (see MNL 60:33-34).
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
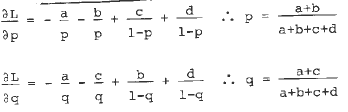{kind=link}