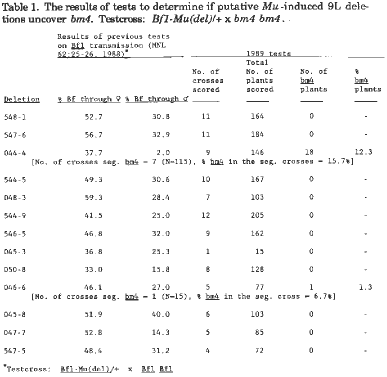
Thirteen of these putative deletion stocks were tested to determine if they uncovered the bm4 locus, which is distal to Bf1. Bf1 seedlings (putative genotype = Bf1-Mu(del)/Bf1) from the Bf1-Mu(del)/+ x Bf1 Bf1 crosses reported in the 1988 News Letter (Tables 1 & 2) were transplanted to the field, and the mature plants were pollinated by pollen from mature bm4 plants. If a putative deletion extends as far as the bm4 locus, brown midrib plants should be observed in the progeny of the cross in approximately the frequency reported for Bf1 seedlings in the female crosses in the 1988 article.
Only two of the 13 putative deletions gave results that suggested that bm4 might be uncovered by the deletion (i.e., 044-4, 046-6, Table 1). However, the frequency of bm4 plants is much below the expected frequency. Of the 9 crosses involving plants heterozygous for the putative 044-4 deletion, two did not segregate for bm4 plants. The percentage of bm4 plants in the 7 progeny that did segregate was 15.7%, which is considerably less than the 37.7% female transmission observed in the 1988 tests. The heterozygous deletion 044-4 plant had semisterile pollen in which the abortive grains were partly filled with starch. Pollen for 15 of the 18 segregating bm4 plants was scored and each had semisterile pollen with abortive partly filled pollen grains. In addition, all segregating brown midrib plants were shorter than their green siblings. Thus, the bm4 plants would seem to be heterozygous for a deletion that involves the Bf1 and bm4 loci. But why the reduced frequency of the deletion plants in these crosses? Perhaps the hypoploid Bf1 bm4 condition is not fully viable. If so there should be a reduced stand in these 9 progenies. From each cross 20 kernels were planted. For the 9 crosses of deletion 044-4 scored, the following numbers of plants were observed with the number of bm4 indicated in parentheses: 12(1), 15 (2), 14, 15 (1), 19 (2), 17 (4), 19 (5), 18 (3), 17. If all the missing plants are assumed to be Bf1 bm4 hypoploids that did not survive (an assumption that very likely is not true), the frequency of hypoploids (heterozygous deletion plants) would be 28.9%, a percentage of transmission 8.8% less than the 37.7% transmission frequency observed in 1988 when plants carrying this deletion were crossed as females. However, because some of the plants that did not survive in these families probably were normal, 28.9% is likely an over-estimate of the frequency of hypoploid Bf1 bm4 kernels produced by this deletion.
Table 1. The results of tests to determine if putative Mu-induced 9L deletions uncover bm4. Testcross: Bf1-Mu(del)/+ x bm4 bm4.
In the five families of deletion 046-6 grown, only one short bm4 plant was found (1.30%). If it is again assumed that the kernels that did not produce plants were hypoploid for a Bf1 bm4 deletion, the percentage of Bf1-Mu(del)/bm4 kernels would be 24.0%, which is far short of 46.1% female transmission frequency of this deletion in the 1988 tests.
Why the lower than expected frequency of bm4 plants in the crosses with these two deletions? Perhaps failure of all Bf1-Mu(del)/bm4 kernels to germinate or the inability of plants of that genotype to survive can explain the deficiency for deletion 044-4. In this case, the deficiency could be due to chance alone. In the case of deletion 046-6, however, such an explanation seems unlikely. Perhaps the one bm4 plant observed was due to a mutation of the wild type allele in what would otherwise have been a +/bm4 sibling, or a loss of the chromosome 9 homologue with the wild type allele, or at least a segment of it involving the allele in such a plant. The bm4 plant had normal pollen, which is not expected if this plant was monosomic for chromosome 9 or heterozygous for a deletion involving Bf1 and bm4. Previous work with reciprocal translocations has shown that plants heterozygous for terminal Bf1-bm4 deletions as a result of the functioning of duplicate-deficient elements of reciprocal translocations in eggs have semi-sterile pollen, with the abortive grains being partly filled (similar to that observed for deletion 044-4). Thus, it is unlikely that the bm4 plant observed in the crosses of deletion 046-6 is due to hypoploidy for this deletion.
In summary, out of 13 putative deletions involving Bf1, only one seems to involve the bm4 locus. Thus most of the remaining 12 are probably not terminal deletions. The 044-4 deletion could be terminal, but just because it seems to include the bm4 locus does not necessarily mean that it does.
In 1988, 10 kernels each from crosses of 100 different putative deletion events involving the Bf1 locus were sown. In each cross, half of the kernels would be expected to be of the putative genotype Bf1-Mu(del)/+ and half Bf1/+. All plants were self-pollinated and scored for the segregation of Bf1 seedlings. The Bf1/+ plants would segregate 3 normal to 1 Bf1 seedlings, while heterozygous deletion plants, if they exist, might have a reduced Bf1 frequency if the deletion is not completely selected against through the pollen, or no Bf1 seedlings at all if there is very rare or no male transmission. Some of the Mu-induced Bf1 events are likely to be Bf1 mutations. In families with such mutations, all plants would segregate 3 wildtype:1 Bf1. Some of the seedlings selected from the original screening as ones that carried Mu-induced events at the Bf1 locus were of doubtful classification. They had phenotypes such as weak zebra patterns of blue fluorescence or sectors of blue fluorescent tissue. If the phenotypes of such plants were not due to genetic events involving the Bf1 gene (pseudo Bf1 event), some of the outcross plants that were self-pollinated also would not segregate for Bf1. In summary, all families would be expected to have half of the selfed progenies segregating 3:1 for Bf1 seedlings. For the other half of the selfed progenies there would be expected to be families which segregated 3:1 for Bf1 (putative small deletions or Bf1 mutations), less than 25% Bf1 seedlings but not 0% (putative deletions with poor pollen transmission), or no Bf1 seedlings segregating (putative deletions not transmitted through the pollen, or pseudo Bf1 events). Table 2 summarizes the results of these tests.
Table 2. Summary of the results of the selfing tests of stocks carrying putative Mu-induced deletions of the Bf1 region of chromosome 9.*
|
|
|
|
|
|
|
|
|
|
|
|
|
|
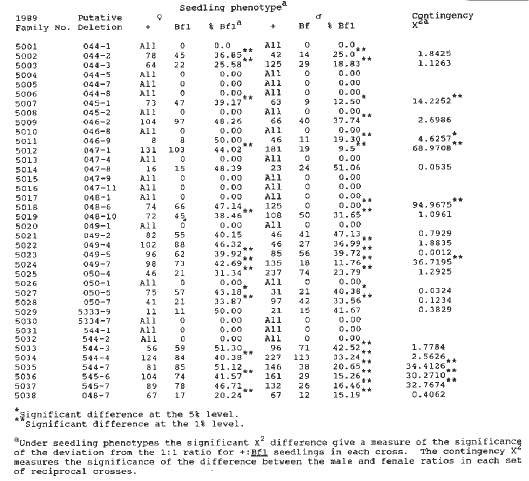
Last summer, 10 kernels from each of 38 selfed ears that did not segregate for Bf1 seedlings were sown and the resulting plants reciprocally crossed to Bf1 Bf1 stocks (Table 3). Fifteen of the original Bf1 events turned out not to be transmitted (i.e., no Bf1 seedlings were observed in these tests). Some of these were expected because the classification of the original Bf1 seedling was doubtful (e.g., pseudo Bf1). Most, however, were originally classified as good Bf1 seedlings. The question is why Bf1 is now lost. At least three explanations are possible: 1) The original mutational event was an early somatic event that did not involve the ear lineage. 2) Methylation of the element at the Bf1 locus has occurred and the mutant phenotype is not observed (See Martienssen et al., EMBO J. 8:1633-1639, 1989, for a similar phenomenon with a Mu-induced mutant of hcf-106.) 3) The transmission or viability of the original mutant is so poor that it has been lost. Twelve of the mutants have equal frequencies of Bf1 seedlings in the male and female outcrosses. Only three of these have 1:1 ratios (047-8, 049-2, 5333-9). The rest have equal male and female transmission frequencies but with less than 50% Bf1 seedlings (e.g., 049-5 with a Bf1 frequency of 39.92% in the female cross and 39.72% in the male cross). There is no indication of deletions being involved in these lines. These mutant alleles, however, do seem to either have lowered male and female transmission or are partially lethal in zygote or embryo. Three mutants (046-2, 049-4, 544-3) had good 1:1 ratios when crossed as females and significantly less than 1:1 ratios when crossed as males. Yet the frequencies of Bf1 seedlings in female and male crosses were not significantly different. A very small deletion may be involved in these. Eight of the mutants have frequencies of Bf1 seedlings in the male cross that are significantly less than 50% Bf1 and also have frequencies of Bf1 seedlings in the male cross that are significantly less than those in the female crosses. These are likely candidates for being deletions. Three of these (045-1, 049-7, 545-6) seem to have reduced female transmission as well because the frequencies of Bf1 seedlings in the female crosses are significantly less than 50%. These three could be larger deletions that are reduced in both male and female transmission, while the remaining 5 are smaller deletions with only reduced male transmission.
Table 3. Reciprocal crosses of plants from the selfed progeny of heterozygous plants with putative Mu-induced Bf1 deletions.
One interesting fact should not be lost
sight of in these tests. The material grown for these reciprocal tests
came from selfed ears, and the seedling tests of the kernels from these
ears did not segregate for Bf1 seedlings. Yet in many of the reciprocal
crosses to homozygous Bf1 stocks, significant numbers of Bf1
seedlings are observed. In all families where Bf1 seedlings were
observed, some of the reciprocal crosses gave no Bf1 seedlings,
as would be expected because the plants that were selfed were heterozygous
for the wildtype allele at the Bf1 locus. In all the reciprocal
tests only one was found that had all Bf1 seedlings expected of
a homozygous, Bf1-Mu plant. Thus, in spite of the fact that the
reciprocal tests indicate, for most of the crosses in Table 3, that Bf1
is readily transmissible through both male and female, homozygous Bf1
parental plants are not or are only very rarely observed. Why? For those
cases where there are indications that a deletion is involved, it is very
likely the deletion is lethal in the homozygous condition. In the other
cases, the Mu-induced events involving the Bf1 region of
the chromosome might result in zygotic or kernel lethals.
Return to the MNL 64 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}