--Barbara Kloeckener-Gruissem and Michael Freeling
Germinal revertants of the Mu3-induced Adh1 mutant 3F1124 (C-H. Chen et al., Genetics 116:469-477, 1987) were isolated. Homozygous mutant plants were grown in isolation from Adh1-3F progenitor plants; these plants were selfed or sib-crossed. Approximately 30,000 resulting kernels were germinated in aerated water for 7 days. Only revertants with partial or fully reverted ADH activity can germinate under these conditions.
Six revertants were recovered that express about 100% ADH1 activity in the scutellum. DNA sequences (100bp) surrounding the Mu3 insertion site of 2 of the 6 revertants were amplified in the polymerase chain reaction (PCR), cloned and sequenced. The DNA sequence of both revertants is indistinguishable from the Adh1-3F progenitor sequence. Yet another revertant, Adh1-3F1124r53 (see B. DeFrancisci et al. this issue), was also found to have no Ac/Ds-typical footprint left upon excision of Mu3.
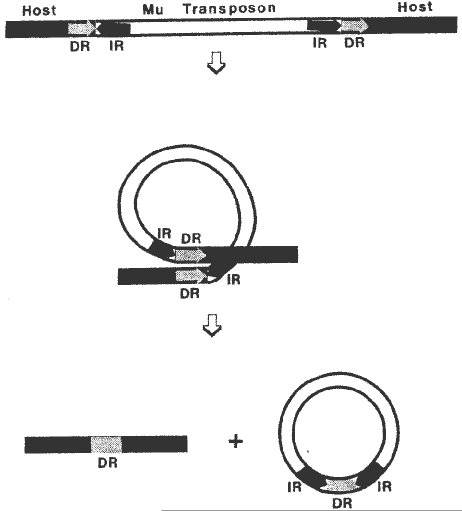
One possible mechanism to explain clean Mu3 excision is a recombination event similar to phage lambda excision, and is illustrated in Figure 1. DNA folds to achieve alignment between the 9bp direct repeats (that resulted from the duplication of host DNA upon insertion of the element). Stabilizing proteins are most likely required to keep the 200bp inverted repeats at the ends of the element from aligning to each other as well as to hold the "loop-structure" in place. A recombination enzyme (not necessarily an enzyme encoded by a Mutator element) may catalyze the cutting and rejoining of the ends yielding the chromosomal copy which is identical to the Adh1-3F allele and a hypothetical circular molecule of the Mu3 element that carries the 9bps from the host DNA duplication between the terminal inverted repeats of Mu3. Thus, the exact sequence of the progenitor allele--shown in the lower left corner of Figure 1 as carrying its single, normal direct repeat (DR)--is restored after an excision. A population of Mu transposon circles of varying sizes has been identified previously by Sundaresan and Freeling (Proc. Natl. Acad. Sci. 84:4924-4928). We are now looking for the DNA sequence between the inverted repeats using the PCR. We predict that each Mu circle will carry a different 9bp insert.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}